网信彩票
-
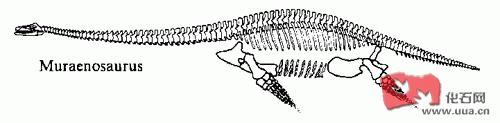
海鱷和蛇頸龍
海鰻龍屬Muraenosaurus地蜥鱷類英國東部牛津粘土一向盛產海生爬行類化石 , 動物群包括蛇頸龍類, 海鱷類及魚龍類��。蛇頸龍類包括長脖子的潛隱龍屬Cryptodidus , 海鰻龍屬Muraenosaurus, Tricleidus , 短脖
海鰻龍屬Muraenosaurus
地蜥鱷類
英國東部牛津粘土一向盛產海生爬行類化石 , 動物群包括蛇頸龍類, 海鱷類及魚龍類����。蛇頸龍類包括長脖子的潛隱龍屬Cryptodidus , 海鰻龍屬Muraenosaurus, Tricleidus , 短脖子的泥泳龍屬Peloneustes , 滑齒龍屬 , 西蒙斯特上龍屬Simolestes及近來發現的厚喙龍屬Pachycostasaurus�����。當地的海鱷類包括地蜥鱷屬及狹蜥鱷屬 , 而魚龍類隻有大眼魚龍一種����。
順道一說,以下的頜骨最初是被L. B. Tarlo (1959)歸屬為上龍類中的Stretosaurus macromerus , 因為L. B. Tarlo 認為這上龍有獨一無二的肩胛骨(scapula)形態 ���。其後於1989年又被Halstead劃歸作滑齒屬 , Noe et al. (2004)認為根據其頜骨形態劃歸作滑齒龍屬也不正確 , 因此把其劃作上龍屬Pliosesaurus , 巨上龍Pliosesaurus macromerus的下頜聯合部分較滑齒龍屬的要長 , 化石收藏牛津大學自然曆史博物館,估計長度為3公尺����。
發現存有咬痕的化石樣本被命為R54(B) , 化石樣本是被劃歸作潛隱龍屬中的Cryptoclidus eurymerus , 樣本保存的也算完整, , 於一些骨骼存有咬痕���。透過這化石上咬痕的研究,從而知道當時海鱷類的習性��。
咬痕的位置顯示隻局限於吃腐肉上,因為屍體可能橫躺在柔軟的海床沉積 , 大多數咬痕是在椎體關節邊緣 , 顯示進食者是於椎間非常小心進食而避免牙齒因顎部施力受損壞 ���。通過比較 , 第二十塊頸椎的齒痕和海生鱷類地蜥鱷屬Metriorhynchus牙齒脊紋非常吻合 , 尺寸比例還是一致 ����。地蜥鱷屬牙齒側部扁平 , 椎體出現的咬痕細小並和對側化石元件咬痕一樣���。根據椎骨上留下的咬痕尺寸及條紋可排除一些大型掠食者所為,例如滑齒龍屬的咬痕間隔較寬及粗糙 , 西蒙斯特上龍屬Simolestes咬痕較輕微及較傾斜�����。也沒有證據顯示那些化石上的紋狀和地殼變動時把屍體埋葬有關 , 例如因埋葬時施壓於骨骼做成的紋痕 , 但這點也不能完全被排除 ��。一些化石上的條紋也部分地和因埋葬屍體時磨擦骨骼導致的相符合 , 但其他的如肋骨上的痕紋卻和這點完全不符��。Hua (1994)指出地蜥鱷類隻是以淺海的頭足類為食,而且隻是根據過去單一的數據進行分析����。Martill (1986b)指出地蜥鱷類胃部樣本包括箭石並把其形容作機會主義的捕食者 , 於他的報導甚至把胃部含有的翼龍骨骼當作魚骨來描述��。根據地蜥鱷屬牙齒的兩種不同的形態:刺穿式及切斷式,可從種間的食性形態學及個體發育年齡的不同 , 反映出它們的進食習性不同之處���。M. superdlosus 及M. leedsi就是具長吻的 , 短吻地蜥鱷M. brachyrhynchus 幼鱷牙齒相對成鱷,輪廓較不扁平壓縮 , 從潛隱龍化石上的咬痕卻和成年短吻地蜥鱷M. brachyrhynchus相吻合����。Martill et al. (1992)指出牛津粘土出土的海生鱷類能披露當時海洋表麵水溫大約維持19-22°C , 而牛津粘土底層顯示深海的水溫維持約15°C , 這顯示海鱷類會偶然下潛至深海因為海洋底部有豐富的食物包括大型海生爬行類的屍體��。潛隱龍的化石樣本顯示地蜥鱷類是進食海床上的屍體 , 並顯示了地蜥鱷類的覓食範圍不隻是在海洋的表麵,也顯示它們是有高機動性的生物 , 能探索廣範的食源及生活環境��。
在本文為网信彩票論壇精華帖���,由網友景德泰嵐提供���,僅供大家學習參考����。
編輯��:劉琮瀅
2022-12-04
-

滄海之牙——(四)��、(五)
根據一些資料顯示 , 從1580年至2008年間襲擊人最多的十大鯊魚中 , 大白鯊是首居第一 , 攻擊人的記錄達377次����,致死人數有205人居氏鼬鯊是居第二 , 攻擊人的記錄達137次����,致死人數有82人 公牛鯊攻擊人
根據一些資料顯示 , 從1580年至2008年間襲擊人最多的十大鯊魚中 , 大白鯊是首居第一 , 攻擊人的記錄達377次���,致死人數有205人;居氏鼬鯊是居第二 , 攻擊人的記錄達137次���,致死人數有82人 ; 公牛鯊攻擊人的記錄有90次����,致死人數有64人����。在第二次世界大戰間 , Nova Scotia號輪船在南非附近被德軍潛艇用魚雷擊沉 , 1000名乘客隻餘192名生還者 ; 1945年7月30日����,有最少60至80名水手被鯊魚襲擊至死 , 二次鯊類襲擊人類的事件都被認為和白鰭真鯊有關 ����。
真鯊科雖沒有鼠鯊科般具有巨大的體型,但它們的凶猛程度絕不亞於鼠鯊科 , 盡管真鯊目也沒有像鼠鯊目般有過稱雄第三紀海洋的光輝歲月 , 但它們經曆漫長的歲月還能維持種類的多樣性 , 而且也有新屬種被發現 , 例如光齒真鯊 , 南非副沙條鯊(Paragaleus leucolomatus)及澳大利亞半沙條鯊(Hemigaleus australiensis)就是真鯊目的新種�����。 相反鼠鯊目像黃昏的太陽 , 經曆了第三紀燦爛光輝的黃金歲月後 , 支持至現在不過隻有16種 , 從多樣性來看二者形成了強烈的對比���。每人對強者的理解各有不同 , 有人會認為有過稱雄海洋的鼠鯊科及黑鯨類是強者 , 但也有人會認為幸存才是強者 , 現存真鯊目中有不少成員早於第三紀時期已存在於地球上 ,有些種類甚至比巨齒鯊還要早出現 , 大西洋斜鋸牙鯊於始新世時期己存在了���。真鯊目成員遍布淺海 , 遠洋 , 深海 , 瀉湖 ,紅樹林,珊瑚礁甚至是淡水 , 它們有底棲性 , 大洋性及淡水性等各種形式, 例如貓鯊屬種就是偏好多沙海床上的底棲性鯊類 ; 東太平洋就能看到數量上百的路氏雙髻鯊在成群遊動 , 大青鯊在大西洋隨強大的洋流追捕魚類及烏賊 ;黑邊擬皺唇鯊(Pseudotriakis acrages)及小齒擬皺唇鯊(Pseudotriakis microdon)就能在1500公尺的水深生活 ; 檸檬鯊就是在紅樹林及瀉湖繁殖的 ; 公牛鯊的調節鹽度能力比任何鯊魚都出色 , 它們會定期出現在河流及湖泊 , 在密西西比河及亞馬遜河中 , 兩處的公牛鯊更能逆流超過2500公厘到達目的地 , 此外巴基斯坦 ,印度及澳大利亞的河流也有露齒鯊屬在那裏長期生活 �����。隻要是有水的地方 ,就沒什麽是它們不能征服的 , 能幸存下來並發展出多樣性的種類 , 真鯊目才是海洋中真正的強者!
真鯊目是起源於晚侏羅紀時期 , 這和鼠鯊目起源的時間相符 , 最早期的鼠鯊目成員是古錐齒鯊屬Paleocarcharias���。像翼手須鯊屬Pteroscyllium牙齒同時具備鼠鯊科及貓鯊科的特征 , 它們是生活於淺海瀉湖的鯊類���。斯皮頓翼手須鯊Pteroscyllium speetonensis 牙齒是被發現於英國斯皮頓阿普第階(aptian)地層 , 牙齒高度達3.5mm , 斯皮頓翼手須鯊Pteroscyllium speetonensis牙齒形狀像錐齒鯊類,有直立的主齒尖及一對發展完善的側齒尖 , 有“V”形的牙根 , 斯皮頓翼手須鯊Pteroscyllium speetonensis有一定的異型齒係 , 細小的牙齒有較少量的唇麵脊 , 後牙有不對稱的牙根裂(基部)及短小的主齒尖 , 後牙有直立的主齒尖及一對等尺寸的側齒尖並和主齒尖相平行���。翼手須鯊屬Pteroscyllium除了斯皮頓翼手須鯊Pteroscyllium speetonensis,還有英國斯皮頓阿普第階(aptian)地層發現的Pteroscyllium ornatum , 英國晚白堊紀科尼亞克階(Coniacian)至坎帕階(Campanian)地層發現的Pteroscyllium hermani , 桑托階(Santonian) 黎巴嫩的Pteroscyllium signeuxi, 德國森諾曼階地層發現的Pteroscyllium nolfi及麥斯特裏希特階(Maastrichtian)摩洛哥的Pteroscyllium lamranii ����。英國晚白堊紀科尼亞克階(Coniacian)至坎帕階(Campanian)地層發現的Pteroscyllium hermani和德國森諾曼階地層發現的Pteroscyllium nolfi關係較密切 , 有細長的牙齒,纖弱的側齒尖及 不規則的紋狀,牙齒唇麵長有直脊; Pteroscyllium ornatum於翼手須鯊屬種中有較大的牙齒 , 牙齒高度能達4.5 mm , 有粗狀及鋒利的主齒尖 , 牙齒主齒尖及側齒尖也長有鋸齒切邊��。 翼手須鯊屬Pteroscyllium和其他貓鯊科的關係至今不明 , 翼手須鯊屬Pteroscyllium牙齒有血管並有著其他貓鯊科的齒係模式 , 但它們的牙齒形態卻像鼠鯊目 , 翼手須鯊屬Pteroscyllium的發現顯示了鼠鯊目和真鯊目於進化枝上是屬於姐妹群���。
德國晚森諾曼階地層發現的鯊類牙齒化石 :
1. Ptychodus decurrens; 2. Chiloscyllium greenei; 3. Lamna appendiculata; 4. Lamna arcuata; 5-6. Isurus denticulatus ; 7. Scyliorhinus aff. destombesi ; 8. Pteroscyllium nolfi; 9. Squalicorax falcatus; 10. Heterodontus canaliculatus; 11. Hexanchus microdon.
貓鯊科(Scyliorhinidae)是真鯊目中最古老的成員 , 已知最早期的成員來自晚侏羅紀時期包括法國的櫛貓鯊Ctenascyllium kimmeridgienus及德國的Macrourogaleus hassei , Macrourogaleus hassei有單一的背鰭,背鰭及臀鰭的位置及大小也不相同, Macrourogaleus hassei有較長的身軀���。科律司鯊Corysodon ceirinensis是另一種發現於德國西南部的鯊類,它和古須鯊屬Palaeoscyllium有著相似位置及大小的背鰭, 科律司鯊Corysodon ceirinensis化石保存的相當清晰可見, 科律司鯊Corysodon ceirinensis曾被認為和古貓鯊屬Palaeoscyllium是同義但至今分類位置還是不明的���。相比科律司鯊Corysodon ceirinensis, 古須鯊屬Palaeoscyllium formosum 的第二背鰭要比前背鰭的小, 腹鰭及臀鰭位置也各不相同, 古須鯊屬Palaeoscyllium formosum牙齒具備精巧脆弱的中齒尖,側齒尖具有許多雕紋����。古須鯊屬的Palaeoscyllium formosum分布於德國及西班牙, Palaeoscyllium formosum的相關近親有英國侏羅紀中期巴通階(Bathonian)地層發現的Palaeoscyllium tenuidens , Palaeoscyllium tenuidens是最古老的古須鯊屬種,於早白堊紀阿爾布階(Albian) 的英國還有Palaeoscyllium reticularis, 不過Palaeoscyllium reticularis則可能是劃作別屬Parasymbolus�����。英國晚白堊紀科尼亞克階(Coniacian)至坎帕階(Campanian)地層發現的Palaeoscyllium striatum 前頜的側齒較大並彎曲及有較細小的側齒尖 , 齒冠基部長有脊紋��。
中生代的英國對真鯊目來說可謂是流奶與蜜之地 , 在當地也發現了許多的真鯊目成員的牙齒化石 , 在這貼會首先論述英國的真鯊目屬種��。
於英國中部及南部侏羅紀巴通階(Bathonian)沉積層發現了數目眾多的鯊魚牙齒化石 , 牙齒化石多達8000顆 , 當地的鯊類超過25種 , 其中有不少是屬於真鯊類 , 發現的軟骨魚類品種如下: 古須鯊屬種 Palaeoscyllium tenuidens., Praeproscyllium oxoniensis., Eypea leesi., 原虎鯊屬種 Proheterodontus sylvestris., 副異齒鯊屬種 Paracestracion bellis., 古長須鯊屬種 Palaeobrachaelurus mussetti., Heterophorcynus microdon., Dorsetoscyllium terraefullonicum., Ornatoscyllium freemani., 偽南喙哈那鯊屬種 Pseudonotidanus semirugosus, Synechodus duffini, Protospinax magnus , Protospinax bilobatus., Protospinax carvalhoi., Belemnobatis kermacki , Belemnobatis stahli , Spathobatis delsatei , Paranotidanus, 弓鯊屬種Hybodus levis及 Breviacanthus brevis等等 , 這些軟骨魚類各自局限於不同的生態環境 , 當時的英國正是被一片淺海所覆蓋���。
在當地較常見的真鯊目成員是Eypea leesi , 它的牙齒非常普遍 , 有較大的牙齒����。Eypea leesi牙齒化石被發現於混濁的離岸相環境及碳酸石灰岩 , Eypea leesi牙齒化石於瀉湖及淺水沉積層是沒有發現的 , 這顯示Eypea leesi是一種偏好深海的鯊類����。Eypea leesi的後齒有較低的齒尖 , 這顯示它是一類偏好以甲殼類為食的真鯊類���。另一種真鯊目成員古須鯊屬種Palaeoscyllium tenuidens則和Eypea leesi不相同 , Palaeoscyllium tenuidens則是較偏好淺水的真鯊類���。當地的真鯊目成員還有Praeproscyllium oxoniensi牙齒化石於當地的瀉湖沉積層非常常見 ,那裏的鹽度較低 , Praeproscyllium oxoniens是非常偏好淺水的鯊類 ����。於當地發現的Dorsetoscyllium terraefullonicum, Ornatoscyllium freemani, Heterophorcynus microdon及古長須鯊屬種Palaeobrachaelurus mussetti是屬於須鯊目成員 ,四種須鯊目成員都是較偏好淺水的鯊類�����。
於英國晚白堊紀科尼亞克階(Coniacian)至坎帕階(Campanian)地層也出土了大量鯊類及鰩類的牙齒化石 , 其中有不少是屬於真鯊目成員包括偽貓鯊屬種Pseudoscyliorhinus reussi , 厚齒貓鯊Crassescyliorhinus germanicus , 貓鯊屬種Scyliorhinus elongatus, Scyliorhinus brumarivulensis., 古貓鯊屬種Palaeoscyllium sp., 原寬瓣鯊Prohaploblepharus riegrafi, 櫛貓鯊屬種Cretascyliorhinus sp., 二個貓鯊科不定種(Scyliorhinidae incertae sedis), 翼手須鯊屬種Pteroscyllium hermani , 原貓鯊屬未定種Protoscyliorhinus sp., 白堊須狗鮫Leptocharias cretaceous , Palaeogaleus havreensis , 副皺唇鯊屬種Paratriakis subserratus , Paratriakis tenuis sp , Paratriakis sp. indet. 及彎齒鯊屬未定種(Loxodon sp) �����。這些白堊紀鯊魚屬種分別歸屬原鯊科(Proscylliidae) , 須狗鮫科(Leptochariidae)及真鯊科(Carcharhinidae) , 顯示真鯊目成員於當地是非常繁盛��。
偽貓鯊屬Pseudoscyliorhinus屬種包括森諾曼階(Cenomanian)德國的Pseudoscyliorhinus schwarzhansi及桑托階(Santonian)至坎帕階(Campanian)英國的Pseudoscyliorhinus reussi�����。Pseudoscyliorhinus reussi 有較平坦的齒根及少量的營養構 , 前齒唇麵基部處有網狀的的紋�����。偽貓鯊屬Pseudoscyliorhinus的牙齒和其他貓鯊科屬種很不相同,但是Pseudoscyliorhinus reussi曾被劃作貓鯊屬種Scyliorhinus reussi ����。現存的貓鯊屬種幼體有較大的牙根 , 幼貓鯊屬牙根也較成體的來得平坦 , 因此發現的Pseudoscyliorhinus reussi牙齒化石也可能是來自幼鯊的 , 發現的偽貓鯊屬Pseudoscyliorhinus reussi牙齒化石寬度僅隻有3毫米���。
Crassescyliorhinus學名有厚的意思 , 故名思義 , Crassescyliorhinus牙齒較其他貓鯊科成員的牙齒來得厚,因此學名也可翻譯作厚齒貓鯊���。厚齒貓鯊屬Crassescyliorhinus germanicus有非常對稱的粗實牙齒 , 中央的主齒尖呈圓錐形並占了主齒冠很大的麵積比例 , 牙根沒有營養構 , 齒冠表麵存有脊紋���。厚齒貓鯊Crassescyliorhinus germanicus和桑托階(Santonian)至馬斯特裏赫特期(Maastrichtian)貓鯊屬種Scyliorhinus musteliformis關係密切 , Scyliorhinus musteliformis可能也是歸屬厚齒貓鯊屬,如果真的劃歸的話,那厚齒貓鯊屬Crassescyliorhinus則一直延存至晚白堊紀的馬斯特裏赫特期(Maastrichtian)���。
英國東部白堊紀阿爾必階(Albian)晚期的Hunstanton Formation發現了另一類貓鯊科屬種Parasymbolus reticularis的牙齒化石 , 被發現一共有五塊牙齒化石 , Parasymbolus reticularis齒冠唇麵上長有網狀的紋��。Parasymbolus屬的主模式種是晚侏羅紀法國的Parasymbolus octevillensis , Parasymbolus octevillensis及Parasymbolus reticularis齒冠及牙根形態相似 ,齒冠唇麵長有鋒利的脊 , 但不同在於Parasymbolus reticularis齒冠唇麵長有網狀的紋 , Parasymbolus reticularis牙根的營養構和Parasymbolus octevillensis幼鯊的相似 ���。Parasymbolus reticularis的發現顯示了Parasymbolus是很長壽的屬 , 屬種生存年代是侏羅紀早期至白堊紀早期���。
英國晚白堊紀科尼亞克階(Coniacian)至坎帕階(Campanian)發現的原寬瓣鯊屬種(Prohaploblepharus sp)的牙齒就非常類似現在的埃氏寬瓣鯊(Haploblepharus edwardsii) , 不過原寬瓣鯊屬種(Prohaploblepharus sp)有較直的主齒尖,齒尖高度值要較寬度值來得大 , 且長有兩對分散的側齒尖 , 齒冠基處也長有縱向的脊紋����。原寬瓣鯊屬(Prohaploblepharus)的主模式種是坎帕階(Campanian)德國的Prohaploblepharus riegrafi , 牙齒的紋和英國晚白堊紀科尼亞克階(Coniacian)至坎帕階(Campanian)地層發現的史前貓鯊屬種Scyliorhinus brumarivulensis非常相似 , 不過原寬瓣鯊屬種Prohaploblepharus riegrafi有較大的牙齒 , 牙根也有所不同�����。
和鼠鯊目相反 , 貓鯊科於始新世的普裏阿邦階(Priabonian )的烏拉爾山脈隻是屬於地域性 , 當地主要的貓鯊科隻有三種 , 包括Premontreia uralica , Foumtizia zhelezkoi 及Foumtizia pattersoni ���。Foumtizia屬的主模式種是Foumtizia abdouni , 這屬是小型貓鯊科屬係,它們的牙齒相當細小但具有尖銳的齒冠 , 有發展完善的突出齒尖 , 齒冠唇麵下部膨大並被琺琅質覆蓋及懸垂於齒根之上 , 齒根有發育良好的營養構���。Foumtizia abdouni生活於古新世塔內提階(Thanetian)的摩洛哥 , 其它屬種包括古新世達寧階(Danian)摩洛哥的Foumtizia gadaensis 及Foumtizia arba ,始新世伊普雷斯階(Ypresian)英國的Foumtizia pattersoni����。於西伯利亞西部出土了無數鯊類的牙齒 , 其中也有真鯊目中的貓鯊科牙齒化石發現 , Premontreia uralica., Foumtizia zhelezkoi 及Foumtizia pattersoni 就是被發現於西伯利亞西部盆地的三種貓鯊科成員 , 三種貓鯊科成員牙齒化石的發現顯示了它們是於晚始新世時期海洋退減時停留在Peri-Tethys,該地北部於普裏阿邦階(Priabonian)氣候冷卻作用促進了當地物種形成���。
Premontreia是一類已滅絕的貓鯊科屬種 , 大多是生存於早古新世至早始新世 , 它們有相對較大的齒尖 , 高度能達6 mm , 齒舌麵平滑及外側突起 , 齒唇麵則沒有像舌麵般有較明顯的突麵 , 基部有琺琅脊(背褶疊), 牙齒長有一至二對側齒尖 , 牙齒有膨大的齒根及有顯著直立的齒冠���。Premontreia屬包括達寧階(Danian)至始新世伊普雷斯階(Ypresian)前期摩洛哥的Premontreia subulidens , 晚古新世塔內提階(Thanetian)摩洛哥的Premontreia peypouqueti , 始新世伊普雷斯階(Ypresian)晚期法國的Premontreia degremonti , 始新世普裏阿邦階(Priabonian)俄國的Premontreia uralica����。始新世伊普雷斯階晚期法國的Premontreia degremonti化石是較常見的 , 於維吉尼亞始新世伊普雷斯階晚期的Nanjemoy Formation也發現了Premontreia degremonti相似種Premontreia cf degremonti的牙齒化石 , Premontreia屬種還有古新世塔內提階(Thanetian)的法國北部 , 始新世的比利時 , 法國及美國的Premontreia gilberti及漸新世至上新世歐洲的Premontreia dachiardi��。近來於比利時始新世路德階地層也發現了新種Premontreia lutetiensis , 它的牙齒高度達8毫米 , 它的牙齒是Premontreia屬中最大的 , Premontreia degremonti牙齒高度也不過才隻有6毫米��。
光尾鯊屬(Apristurus)一共至少有25種 , 大多數體型都是非常細小 , 像圓鰭光尾鯊(Apristurus spongiceps)全長才隻有50厘米 , 巴拿馬光尾鯊(Apristurus stenseni)全長隻是超過23厘米 , 但是屬中的駝背光尾鯊(Apristurus gibbosus)雄鯊可達3.88米, 雌鯊更能達4.1米���。一些種類如無斑光尾鯊 Apristurus acanutus甚至能棲息於840米的水深 , 南非光尾鯊(Apristurus saldanha)的棲息深度也能達915米 , 北美的加州光尾鯊(Apristurus kampae)及深水光尾鯊(Apristurus profundorum)更能棲息於1800米水深的環境���。 不像貓鯊屬Scyliorhinus, 光尾鯊屬(Apristurus)的化石記錄相對要稀少得多 , 已知的史前的光尾鯊屬僅隻有法國始新世地層發現的Apristurus sereti , 於意大利Fiumefreddo更新世時期沉積層發現了冰島光尾鯊的親近種Apristurus aff laurussonii的牙齒化石 , 於當地也發現了黑口鋸尾鯊( Galeus melastomus)的牙齒化石���。
鋸尾鯊屬(Galeus)一共約十二種, 於法國的早期中新世地層留下最古老的化石記錄,像現存的黑口鋸尾鯊( Galeus melastomus)於中新世的法國及德國均有化石記錄,已滅絕的成員還有Galeus pristodontus , 不過據說墨西哥始新世中期地層也有鋸尾鯊屬未定種的牙齒化石發現���。梅花鯊屬(Halaelurus)及絨毛鯊屬(Cephaloscyllium)於印太平洋有廣泛的分布 , 其中澳洲也有兩屬的分布 , 梅花鯊屬(Halaelurus)及絨毛鯊屬(Cephaloscyllium)都各有一種生活於東太平洋 , 兩個屬於大西洋都沒有分布���。絨毛鯊是喜歡棲居於海床石縫及大型海藻,受到驚嚇時會吞下水或空氣使身體膨脹到正常身軀的兩倍大左右 ,使敵人難以把它拉離移居點 ���。於加利褔亞Kern County也發現了絨毛鯊屬未定種的化石����。
新軟骨魚類Neoselachians首次於古生代出現 , 包括所有現存的鯊類及鰩類 , 牠們的牙齒由三層琺琅質覆蓋���。除了鯊類和鰩類外 , 還包括馬克默多鯊Mcmurdodus , 吉特鯊Ginteria, 哥利鯊Cooleyella ,Reifia , Hueneichthys, Rhomphaiodon ,小弓鯊"Hybodus" minor, 哥臣鯊Grozonodon candaui , 晚三迭紀至早侏羅紀西歐的Jurabatos cappettai , 早侏羅紀意大利的Ostenoselache stenosoma , 晚白堊紀德國的副扁鯊Parasquatina cappetai及始新世的Odontorhytis 等許多史前軟骨魚類 ; 真正的鯊類除了現存的九個鯊目外 , 還有已滅絕的古鰭棘鯊目(Synechodontiformes) , 古鰭棘鯊目包括原鰭棘鯊屬Protospinax, 楔齒鯊屬Sphenodus, 古鰭棘鯊屬Synechodus, 副楔齒鯊屬Paraorthacodus , Welcommia , 至於偽南喙哈那鯊屬Pseudonotidanus是否歸屬古鰭棘鯊目或者是歸屬六鰓鯊目則尚存疑問 , 於以下並不介紹偽南喙哈那鯊屬Pseudonotidanus���。
最早期已知的新軟骨魚類成員是馬克默多鯊Mcmurdodus , 生存於泥盆紀中期(距今三億九千萬年)的西昆士蘭�����。今人震驚的是 , 這種鯊類較裂口鯊Cladoselache及南極鼠鯊Antarctilamna更符合現在所有鯊目成員的先祖 , 馬克默多鯊Mcmurdodus牙齒琺琅質有現在鯊魚牙齒般特質 , 現存的鯊類有多層的齒排及不同的牙齒形式 , 這特征於大多數遠古鯊類是沒有 ,但異棘鯊類Xenacanthus卻是例如的����。如果馬克默多鯊Mcmurdodu , 裂口鯊Cladoselache及南極鼠鯊Antarctilamna是同一時期的世係成員的話 , 那麽泥盆紀早期已進行了很巨大的軟骨魚類大幅射進化���。盡管裂口鯊Cladoselache , 南極鼠鯊Antarctilamna及其它早期軟骨魚類堅持了一段時間,但隻有馬克默多鯊Mcmurdodus完成了一個偉大的進化夢想����。
馬克默多鯊Mcmurdodus和現在鯊類關係怎樣是相當不明,像大多數早期的鯊類般, 馬克默多鯊Mcmurdodus也僅隻留下少量的牙齒化石, 牠們的下頜齒像現存的六鰓鯊屬和竺鱗鮫屬 , 不過最後的馬克默多鯊Mcmurdodus和最早的六鰓鯊科成員是存有一億九千萬年的空隙 , 但馬克默多鯊Mcmurdodus作為最早的新軟骨魚類成員已是被公認的事實����。
吉特鯊Ginteria fungiforma是生存於石炭紀早期的英國及俄羅斯的板鰓類 , 牠們的牙齒非常細小獨特 , 牙齒最高值隻有0.54 mm , 齒冠像一個台座 ����。吉特鯊Ginteria fungiforma 齒冠底部長有數目眾多的小孔可能是存有單一的血管連接齒冠底部及頂部����。另一類軟骨魚類哥利鯊屬Cooleyella也有類似的牙齒 , 但吉特鯊Ginteria fungiforma的齒冠表麵要較哥利鯊屬Cooleyella更呈凹麵 , 相比較下吉特鯊Ginteria fungiforma牙齒基部表麵較平坦 , 而哥利鯊屬Cooleyella牙齒要較吉特鯊Ginteria fungiforma的大得多���。
Duffin 及Ward 就於英格蘭及威爾士石炭紀前期地層及美國內華逹州二疊紀早期地層發現的牙齒化石建立了一個新科Anachronistidae , 主模式種是Anachronistes fordi ����。但後來Duffin, Richter 及Neis宣稱Anachronistes是和哥利鯊屬Cooleyella為同物異名��。哥利鯊屬種Cooleyella fordi有三十五顆牙齒 , 牙齒長度介乎於1亳米至2亳米 , 牙齒化石是被發現於北韋爾斯的克盧伊達郡 , 牙齒有圓錐形的主齒尖 , 並有發育良好的側齒及基部突緣 , 牙根呈V狀 , 牙根是布滿血管���。哥利鯊屬Cooleyella有著扁鯊屬Squatina及須鯊屬Orectolobus的特征如牙齒根部中央有著布滿血管的小孔 , 正視時哥利鯊屬Cooleyella牙齒排列明顯有數行 , 這點於扁鯊屬Squatina是存在的 , 齒片間的牙齒齒側互相重疊 , 哥利鯊屬Cooleyella牙齒是被認為適合以壓碎甲殼類為食且哥利鯊屬Cooleyella是一類底棲性的軟骨魚類����。Duffin及 Ward認為哥利鯊屬Cooleyella可能像現在的扁鯊屬Squatina及須鯊屬Orectolobus有著較扁平的身體及較大的胸鰭 ���。
新軟骨魚類Neoselachians於三疊紀時期繁洐 , Mucrovenator和相關屬種小弓鯊"Hybodus" minor , Rhomphaiodon是當時的代表 , 牠們的牙齒形態有著晩三疊紀時期的古鰭棘鯊目Synechodontiformes成員特征 , 並可能也歸屬於古鰭棘鯊目Synechodontiformes ���。 古鰭棘鯊目Synechodontiformes 中的古鰭棘鯊屬Synechodus數個種類是生存於三疊紀時期 , 牙齒形態是低且具有較大的齒根 , 齒根布滿不規則的小孔 , 有單一或多直立的尖峰 , 這些特征於其它三疊紀的種類如哥臣鯊Grozonodon及偽姥鯊Pseudocetorhinus是具有, 偽姥鯊Pseudocetorhinus是一種三疊紀的濾食者鯊類��。
盡管這些三疊紀古鰭棘鯊目Synechodontiformes種類顯微結構上有些特征是不相同,但彼此間的關係已是非常密切 ��。1977年發現的Hueneichthyes , 對這種鯊魚的認知還是非常少 , 但於1999年已被Cuny及 Benton視作可能是歸屬古鰭棘鯊目synechodontiform�����。Hueneichthyes是晚三迭紀的軟骨魚 , 但牙齒形態卻有別於其它三迭紀的新軟骨魚類(neoselachians) , Hueneichthyes 有著細長的牙根及少量的小孔 , 平坦的齒冠懸垂於齒根之上���。
1980年發現晚三疊紀的Reifia minuta, 牠的牙齒也是不同其它三疊紀的新軟骨魚類neoselachians , Reifia minuta有著平坦的齒冠及纖細的齒根 , 齒根長有數目較少的小孔�����。形態學上的獨特之處顯示新軟骨魚類Neoselachii於三疊紀中期或早晚三疊紀時期發生輻射性的分枝演化 , 1982年發現的Vallisia 及1978年發現的偽鎧鯊Pseudodalatias 牙齒形態和新軟骨魚類neoselachians一些成員相似但沒有典型的新軟骨魚類neoselachians顯微結構 , 牠們的關係仍是不明的���。
古棘鰭鯊目Synechodontiformes成員有著相同的牙齒形態 , 特點是包括有較低及較大的牙根 , 牙齒有單一及多齒尖 , 牙根上滿布不規則排列的小孔 , 三迭紀的哥臣鯊Grozonodon 及偽姥鯊Pseudocetorhinus 並不屬於古棘鰭鯊目 , 但卻有著以上特征僅隻是牙齒中微觀結構上的分別����。
有關晚三疊紀時期的古鰭棘鯊目Synechodontiformes及六鰓鯊目(Hexanchiformes) , 古鰭棘鯊屬Synechodus及其它古鰭棘鯊類的關係是被視作存有疑問的 , Maisey (1985) , Maisey (2004) 及Cappetta (1987)認為牠們不能形成一個單元的類群����。古鰭棘鯊目Synechodontiformes看似缺少許多起源的先祖特征而且牠們也可能是並係(paraphyletic) , 牠們的齒式許多特征具有和六鰓鯊目相似性 , 例如牙齒上都具有多尖峰 , 有對稱的聯合齒 , 特化具壓碎特征的後外側牙齒 , Welcommia及偽南喙哈那鯊Pseudonotidanus二屬鯊類形態就介乎於古鰭棘鯊目Synechodontiformes及六鰓鯊目之間���。盡管有著這些相似性 , 所有古鰭棘鯊目Synechodontiformes成員有著星狀椎骨(asterospondylous vertebra) , 和六鰓鯊目的單環椎(cyclospondylous vertebra)明顯不同 ��。盡管椎骨上的不同顯示兩個鯊目之間的關係疏遠 , 但中生代的六鰓鯊目Hexanchiformes成員具有星狀椎骨(asterospondylous vertebra)且椎骨是有縱向的薄層 , 這顯示較古鰭棘鯊目Synechodontiformes來得晩出現的六鰓鯊目單環椎結構是具祖先特征並是一種過渡的型態����。
於三疊紀晚期的淺海海床生活了一種小型鯊類---小弓鯊'Hybodus minor , 牠的牙齒是平坦且寬闊,顯示牠們的牙齒是以壓碎食物為主的 ,故名思義牠們是屬於弓鯊科, 但最近對牙齒的研究顯示牠們可能是歸屬新軟骨魚類Neoselachii ���。於發現小弓鯊'Hybodus minor牙齒化石的地方同時也發現了新軟骨魚類(Neoselachii)另一品種Nemacanthus monolifer的棘部 , 這些鯊魚沒有完整化石被發現 , 因此最初也難顯示化石是否屬於同一物種 , 直至小弓鯊有完整的化石被發現才可解答鯊類間的關係��。Nemacanthus monolifer要比古鰭棘鯊屬[Palaeospinax]要來得早 , 牠和古棘鯊屬[Palaeospinax]有密切的關係 , Nemacanthus monolifer也被認為有可能是古棘鯊屬的先祖, Cappetta於1987年把Nemacanthus monolifer劃作古棘鯊科(Palaeospinacidae) , 他們認為Nemacanthus monolifer是真正的新軟骨魚類或者是屬於新軟骨魚類的姐妹群���。
Rhomphaiodon nicolensis被發現於法國三疊紀晚期地層, 牠有百多顆牙齒化石, 從形態上Rhomphaiodon nicolensis牙齒和小弓鯊"Hybodus" minor 的相似 ; 另一種於法國東部Grozon發現的新軟骨魚類哥臣鯊Grozonodon candaui的牙齒琺琅質結構和小弓鯊的相同 , 二者可能存有密切關係���。
Reifia minuta 僅隻有五顆細小的牙齒化石發現 , 化石發現於德國諾利克階(Norian)前期地層 , Duffin (1980)從Reifia minuta 的牙齒判斷牠們和須鯊目(Orectolobiformes)關係較密切 , 但是Cappetta (1987)則為認為Reifia minuta 和真鯊目關係要較密切一些 , 但是也有學者認為Reifia minuta的牙齒和鋸鰩目中Sclerorhynchidae的牙齒相似���。
相對來說大多數新軟骨魚(neoselachian)體型細小 , 長度維持於1米內 , 但牠們的牙齒比弓鯊目來得進步 , 具備了抓捕獵物的能力了 , 例如雷蒂亞古鰭棘鯊Synechodus rhaeticus , 牠的牙齒同時具有抓捕及壓碎的能力 ,不過牠們是傾向以甲殼類為食的����。當地的另一種新軟骨魚(neoselachian)偽姥鯊Pseudocetorhinus pickfordi可能是一種濾食性軟骨魚類 , 牠時常被看作姥鯊科成員 , 但是最早的姥鯊屬是被發現於南極西摩島始新世中期地層��。Hueneichthys costatus, Rhomphaiodon nicolensis,小弓鯊"Hybodus" minor及哥臣鯊Grozonodon candaui則屬於新軟骨魚(neoselachian)最原始的成員並和古鰭棘鯊目Synechodontiformes有密切的關係 , 這些原始的新軟骨魚類和古鰭棘鯊目Synechodontiformes彼此有著相似的齒冠 ,但齒根卻是不相似的 ; 至於Reifia minuta及雷蒂亞古鰭棘鯊Synechodus rhaeticus二者的分類地位是存有疑問的 , 二者可能是代表另一個晚三迭紀的軟骨魚類世係 ���。
於晚三迭紀至早侏羅紀交界時海平麵改變並強裂影響西歐 , 瑞替階(Rhaetian Age)後歐洲中部及北部都被一片淺的陸緣海所覆蓋 , 於當時西歐的一片淺海中 , 新軟骨魚(neoselachian)種類占了50% , 牠們於西歐進行輻射形進化 ���。這時期的西歐存有一些和新軟骨魚(neoselachian)存有關連的短暫軟骨魚類世係例如Vallisia , Doratodus, 偽鎧鯊屬Pseudodalatias, Raineria ���。 偽鎧鯊Pseudodalatias是具有切斷及壓碎食物能力的牙齒 , 這顯示牠們能以相對較大的軟件動物為食 ��。
早侏羅紀意大利的Ostenoselache stenosoma有著許多獨特的特征包括有較長的腦顱 ,沒有背鰭 ,牠的主齒尖和皺鰓鯊屬及一些三迭紀的古鰭棘鯊目成員的相似 ; Ostenoselache stenosoma是沒有衍生形質(autapomorphic characters) , 對於牠的係統學位置也是不明的��。
距馬克默多鯊Mcmurdodus後的一億四千萬年後海洋出現了另一類屬,古鰭棘鯊類Paleospinax(Synechodus) , 目前對牠們的認識僅來自牙齒化石的發現 , 古棘鯊類Paleospinax是生存於二疊紀早期至始新世時期(約二億五千萬年前至六千萬年前) , 但英國及德國僅隻發現了小量完整的古鰭棘鯊類頜骨及椎骨化石樣本����。古鰭棘鯊全長一般小於三呎但和現在的鯊類特征有很多關連 , 例如有較長的吻部,口器已置於頭部的下方,短小的頜骨隻和顱後相連 , 牙齒有較密的琺琅質及發育良好的椎骨��。牠們身體表麵存有鰭棘 , 類似現在的角鯊科����。盡管對古鰭棘鯊類認知還是很少,但牠們已被認為是屬於早期的真正鯊類了�����。
過去認為古鰭棘鯊目synechodontiform 是於中生代初期出現 , 近來IVANOV 於2005年發表於俄羅斯烏拉爾的二迭紀早期地層發現了古鰭棘鯊屬種Synechodus antiquus的牙齒化石 , Synechodus antiquus 的發現顯示古鰭棘鯊目synechodontiform於古生代晚期已存在於地球上���。
早三迭紀土耳其發現的古鰭棘鯊屬種沒被定種 , 晚三迭紀諾利克階加拿大有Synechodus volaticus , Synechodus multinodosus及Synechodus incrementum , 化石都是被發現於英屬哥倫比亞省(British Columbia) , 這也顯示古鰭棘鯊屬於諾利克階北美旁的大西洋取得分布 ���。於晚三迭紀雷蒂亞階(Rhaetian Stage)至古新世達寧階(Danian)歐洲有廣泛的分布 , 第一種出現於西歐是雷蒂亞階(Rhaetian Stage)法國及英國的雷蒂亞古鰭棘鯊Synechodus rhaeticus ���。侏羅紀的屬種有早侏羅紀海塔其階(Hettangian)盧森堡的Synechodus streitzi 及 Synechodus paludinensis ,錫內穆階(Sinemurian)意大利的Synechodus pinnai , 英國的Synechodus priscus , 侏羅紀中期巴通階(Bathonian)英國西南部的Synechodus duffini , Synechodus levis , 卡洛維階(Callovian)玻蘭的Synechodus prorogatus , 晚侏羅紀牛津階(Oxfordian)德國南部的Synechodus riegrafi 及啟莫裏奇階(Kimmeridgian)英國的Synechodus plicatus ���。
早侏羅紀海塔其階(Hettangian)至錫內穆階(Sinemurian) 丹麥,法國及瑞典的Synechodus occultidens , 普連斯巴奇階(Pliensbachian)英國及瑞典的Synechodus enniskilleni , 托阿爾階(Toarcian)英國的Synechodus egertonia (Synechodus smithwoodwardi)牙齒形態有別於其它古鰭棘鯊屬種而劃歸新屬Palidiplospinax , Palidiplospinax和古鰭棘鯊屬的主要不同之處在於其側齒尖之別���。
白堊紀時期古鰭棘鯊屬則向北美及亞洲遷移發展 , 白堊紀屬種包括豪特裏維階(Hauterivian)至阿爾布階(Albian)英法的Synechodus dubrisiensis , 阿爾布階(Albian)至桑托階(Santonian)俄羅斯及哈薩克的Synechodus dispar , 英國及哈薩克的Synechodus nitidus , 阿爾布階(Albian)英法及哈薩克的Synechodus tenuis , 坎帕階(Campanian)美國蒙大納的Synechodus striatus及加拿大的Synechodus turneri , 於澳大利亞也有古鰭棘鯊未定種的化石記錄����。
古鰭棘鯊屬種於晚白堊紀時期衰落 , 但仍有屬種能支持至第三紀 , 比如出現於桑托階(Santonian)俄羅斯的Synechodus hesbayensis , 牠的化石於比利時晚古新世至早始新世地層都有化石記錄, 桑托階(Santonian)哈薩克的Synechodus lerichei 居然也於比利時古新世地層都有化石發現 , 另一屬種Synechodus faxensis於古新世時期丹麥,法國及比利時取得廣泛的分布 , 不過牠們於古新世時期僅隻局促於西歐 , 於早始新世便完全滅絕了���。
原鰭棘鯊屬Protospinax是生存於侏羅紀中期至晚期西歐等地的鯊類 , 牠一般全長5呎 , 和鰩類及角鯊目關係密切 , 屬種包括侏羅紀中期卡洛維階(Callovian)至晚侏羅紀提通階(Tithonian)德國及玻蘭的Protospinax annectans , 晚侏羅紀啟莫裏階(Kimmeridgean)早期英國的Protospinax planus , 侏羅紀中期巴通階(Bathonian)英國西南部則存有三種包括Protospinax bilobatus , Protospinax carvalhoi及Protospinax magnus , 此外於法國也有原鰭棘鯊屬種化石發現��。於德國早提通階地層就發現了完整的原鰭棘鯊屬種骨骼化石 , 牙齒長有卵形的齒冠及突出的低齒尖 , 牙根呈二裂的 , 牠們的牙齒是具有壓碎食物的能力 , 這看似原鰭棘鯊屬Protospinax是傾向以深海底的甲殼類為食 ���。有關原鰭棘鯊屬Protospinax的分類地位是存有爭議的 , 有專家認為牠是最原始的角鯊目成員 , 也有專家認為牠是屬於鰩類���。最古老的角鯊目成員角鋸尾鯊屬種Squalogaleus lochensteinensis及Squalogaleus woodwardi也曾被劃作原鰭棘鯊屬種 , 但後來則被劃歸角鋸尾鯊屬Squalogaleus ���。
楔齒鯊屬Sphenodus (Orthacodus)於侏羅紀的歐洲很常見 , 於歐洲常可找到楔齒鯊屬Sphenodus的主齒尖及椎骨殘骸化石��。楔齒鯊屬Sphenodus的牙齒較適合把獵物撕成碎塊 , 前牙狹長 , 有較高的尖峰並長有明顯的鋸邊 , 頜前端牙齒外形成S形 , 這些牙齒是處於吻邊的並成短且直的折疊�����。牙根基部平坦 , 並沒有任何溝槽 , 牙根和數目眾多的不規則血管貫通 ���。
早侏羅紀屬種有歐洲的Sphenodus alpinus , 瑞士的Sphenodus helviticus , 晚侏羅紀屬種包括歐洲的提通楔齒鯊Sphenodus tithunius , Sphenodus venulosu , 俄羅斯的Sphenodus stschurowskii , 德國南部的長齒楔齒鯊Sphenodus longidens , 梅加楔齒鯊Sphenodus macer 及光齒楔齒鯊Sphenodus nitidus ���。晚侏羅紀時期楔齒鯊屬種於南北半球都有分布 , 牠們是大洋性鯊類並可能會隨季節遷移回遊 �����。
楔齒鯊屬種於白堊紀時期衰落 ,早白堊紀時期僅有Sphenodus salandianus及Sphenodus subaudianus , 晚白堊紀的屬種包括森諾曼階(Cenomanian)的Sphenodus planus 及桑托階(Santonian)的Sphenodus sennessi , 於澳洲昆士蘭白堊紀地層 , 安哥拉坎帕階(Campanian)地層及南極也發現了楔齒鯊屬種化石 , 於南極科尼亞克階(Coniacian)至坎帕階(Campanian)地層發現的楔齒鯊屬種牙齒化石保存不完整 , 但根據牙齒推斷全長至少長達5米��。楔齒鯊屬種於古新世時期完全絕種 , 最後的屬種是丹寧階(Danian)丹麥瑞典的Sphenodus lundgreni �����。
於德國南部發現的長齒楔齒鯊Sphenodus longidens , 梅加楔齒鯊Sphenodus macer 及光齒楔齒鯊Sphenodus nitidus 可帶來不少關於楔齒鯊的習性等資料 , 三種不同的牙齒 , 顯示牠們的進食方式也有所不同��。
梅加楔齒鯊Sphenodus macer有完整的化石被發現 , 牠有較大的胸鰭 , 從梅加楔齒鯊Sphenodus macer盾鱗腹側被沙石磨擦過的痕跡來看 , 梅加楔齒鯊Sphenodus macer是像扁鯊般的底棲性鯊類��。化石樣本SMNS 80142/44是一條長85厘米的梅加楔齒鯊 , 牠的牙齒隻有8.7 mm , 但是於 Schnaitheim 找到的梅加楔齒鯊Sphenodus macer離散牙齒最大的高度值達18 mm , 這顯示化石樣本SMNS 80142/44中的個體是未完全成熟的個體 , 成年的梅加楔齒鯊Sphenodus macer全長應該可達1.7米��。 從牠的牙齒來看主要以抓緊獵物為主的 , 牠可能是靜伏海底泥沙中 , 出奇不意地突襲獵物 ���。
光齒楔齒鯊Sphenodus nitidus的牙齒高度達22.4 mm , 根據牙齒推測光齒楔齒鯊Sphenodus nitidus全長可能超過 2米�����。光齒楔齒鯊Sphenodus nitidus牙齒也同樣長有鋸邊 , 但是光齒楔齒鯊Sphenodus nitidus和長齒楔齒鯊Sphenodus longidens前齒主齒尖彎曲有所不同 , 這也顯示二者的攻擊進食獵物的方式也有所不同 , 光齒楔齒鯊Sphenodus nitidus前齒以捕捉抓緊小型獵物如魚類及頭足類 , 然後把獵物整個吞下 , 從光齒楔齒鯊Sphenodus nitidus和長齒楔齒鯊Sphenodus longidens化石所在來看 , 二者都是屬於大洋性的鯊類 , 牠們是生活於海洋中層及表層的鯊類 , 這點和梅加楔齒鯊Sphenodus macer是完全相反的 ��。
長齒楔齒鯊Sphenodus longidens的牙齒高度達44 mm , 長齒楔齒鯊Sphenodus longidens全長可超過 4米 ���。長齒楔齒鯊Sphenodus longidens牙齒主齒尖長有鋸邊 , 牠可能像現存的大白鯊一樣 , 透過扭動頭部把海生爬行類的肉撕扯下來 , 因此長齒楔齒鯊Sphenodus longidens牙齒主齒尖大多折斷 , 長齒楔齒鯊Sphenodus longidens更曾被看作是應歸屬鼠鯊目��。長齒楔齒鯊Sphenodus longidens於晚侏羅紀的英國及德國 , 晚白堊紀森諾曼階(Cenomanian)至桑托階(Santonian)分布於日本 , 巨大的體型和鋸齒切邊顯示牠可能以小型海生爬行類為食, 恐怕是史上第一種具掠食性的鯊類��。
副楔齒鯊屬Paraorthacodus於侏羅紀世界各地留下許多牙齒和椎骨化石 , 副楔齒鯊屬Paraorthacodus有較大及長長的主齒尖 , 主齒尖及側齒尖都有鋸邊 , 牠們的牙齒是以抓捕獵物為主的���。副楔齒鯊屬從侏羅紀興起 , 至始新世時期才完全滅絕����。
副楔齒鯊屬Paraorthacodus的分布和六鰓鯊目成員Notidanodon都很相似 , 盡管白堊紀時期副楔齒鯊屬種的種類多樣性並不高 , 但卻於兩極氣候和暖及寒冷的地區都有分布 , 晚白堊紀時期副楔齒鯊屬種於南極都有分布���。從副楔齒鯊屬Paraorthacodus的化石形態來看牠們和須鯊目同樣生活於淺海的鯊類 , 以魚類及軟體動物為食的緩慢鯊類���。
副楔齒鯊屬種包括晚侏羅紀德國的Paraorthacodus kruckowi , 啟莫裏階(Kimmeridgean)至提通階(Tithonian)的Paraorthacodus jurensis , 白堊紀 阿爾布階(Albian)英國,法國及俄國的Paraorthacodus recurvus , 坎帕階(Campanian)蒙大納的Paraorthacodus andersoni , 桑托階(Santonian)至坎帕階(Campanian)哈薩克,德國及比利時的Paraorthacodus nerviensis , 晩白堊紀阿根廷的Paraorthacodus patagonicus ,晩白堊紀桑托階(Santonian)至坎帕階(Campanian)南極北部的南極副楔齒鯊Paraorthacodus antarcticus , 麥斯特裏希特階(Maastrichtian)瑞典的Paraorthacodus conicus , 晚白堊紀至古新世新西蘭的Paraorthacodus sulcatus及Paraorthacodus validus , 晚古新世至始新世北美及英國的Paraorthacodus clarkii及始新副楔齒鯊Paraorthacodus eocaenus則為最後的副楔齒鯊屬種
在本文為网信彩票論壇精華帖��,由網友景德泰嵐提供���,僅供大家學習參考����。
編輯��:劉琮瀅
2022-12-03
-
猶太蜥
猶太蜥Judeasaurus tchernovi是被發現於中東晚白堊紀的白堊世中期(Middle Cenomanian)的水生蜥類 , 它的頭骨具有陸上巨蜥類特征例如有寬闊的前頜,上頜前部扁平壓縮 , 但也同時有滄龍類的洐生特征���。Judeasa猶太蜥Judeasaurus tchernovi是被發現於中東晚白堊紀的白堊世中期(Middle Cenomanian)的水生蜥類 , 它的頭骨具有陸上巨蜥類特征例如有寬闊的前頜,上頜前部扁平壓縮 , 但也同時有滄龍類的洐生特征���。Judeasaurus tchernovi 有狹幼及長長的牙根但牙齒沒有吸收孔(resorption pits) , 有些類似Coniasaurus,但猶太蜥Judeasaurus tchernovi齒冠較細小 , 輕微彎曲及呈圓錐型 ���。Coniasaurus則是生活於堪薩斯州晚白堊紀Turonian期(ca. 92.1–91.2 Ma)西部內海的小型海陸兩棲蜥類����。Judeasaurus tchernovi頭骨非常細小,隻有60至70毫米長 , 頭骨較不完整 ,頜骨分別有14至16顆牙齒 , 牙齒緊密排列,沒有折形牙質�����。
Coniasaurus椎骨化石
Haber and Polcyn (2005)根據Judeasaurus tchernovi和其他海生蜥類包括脹蜥Adriosesaurus 及Pontosaurus,基層滄龍類包括特提斯龍Tethysaurus, Haasiasaurus及 大洋龍屬Halisaurus的比較,最後確定Judeasaurus tchernovi是巨蜥下目(Varanoidea)中的新屬種, 前沿融合及圓狀的方骨顯示猶太蜥Judeasaurus tchernovi和滄龍類有關,它和晚白堊紀另一類海生蜥類Dolichosauridae有關係�����。
猶太蜥Judeasaurus tchernovi的化石
在本文為网信彩票論壇精華帖���,由網友景德泰嵐提供���,僅供大家學習參考��。
編輯����:劉琮瀅
2022-12-03
-

歐洲鯊齒鯨科的演化史
晚漸新世至早中新世許多齒鯨類傳統地被劃作鯊齒鯨科(Squalodontidae) , 但是根據MUIZON (1990)的看法 , 隻有始鯊齒鯨屬(Eosqualodon) , 鯊齒鯨屬(Squalodon) , 凱利鯨屬(Kelloggia) , 褔玻斯鯨屬(Pho
晚漸新世至早中新世許多齒鯨類傳統地被劃作鯊齒鯨科(Squalodontidae) , 但是根據MUIZON (1990)的看法 , 隻有始鯊齒鯨屬(Eosqualodon) , 鯊齒鯨屬(Squalodon) , 凱利鯨屬(Kelloggia) , 褔玻斯鯨屬(Phoberodon)及新鯊齒鯨屬(Neosqualodon)是歸屬鯊齒鯨科(Squalodontidae)����。
巴氏凱利鯨(Kelloggia barbarus)被發現在阿拜塞疆晚漸新世地層 , 牠有著長而寬的厚重吻突 , 臼齒和前臼齒同等大小 , 巴氏凱利鯨(Kelloggia barbarus)有短而強狀的前肢����。
極喙褔玻斯鯨(Phoberodon arctirostris)被發現在巴塔哥尼亞早中新世地層 , 牠有近乎完整的骨骼化石發現 , 腦殼的套疊及輪廓和鯊齒鯨屬(Squalodon)相似 , 但是極喙褔玻斯鯨(Phoberodon arctirostris)有較粗狀的顴突 , 牠的齒式要較鯊齒鯨屬(Squalodon)的原始����。
從牙齒來看歐洲的鯊齒鯨科主要有兩個演化世係 , 其中一個是新鯊齒鯨屬(Neosqualodon) , 艾沙新鯊齒鯨(Neosqualodon assenzae)頭骨能長達21吋 , 每邊頜骨有3顆門齒及1顆犬齒 , 下頜有多達26顆頰齒 , 上頜骨有至少七顆後牙具有鋸邊 ���。艾沙新鯊齒鯨(Neosqualodon assenzae)後頰齒保留了三個較大的葉狀前附尖及後附尖 , 主齒尖很難確認 , 新鯊齒鯨屬(Neosqualodon)有原始的齒冠型態及極端的多齒式 , 在鯊齒鯨科(Squalodontidae)裏是相當特化的 , 牠們的牙齒緊密排列並完全沒有牙間隙 , 齒冠相互重疊����。艾沙新鯊齒鯨(Neosqualodon assenzae)腦殼和鯊齒鯨屬相似 , 鼻骨孔後移至上枕突部分並和眶前成角度����。艾沙新鯊齒鯨(Neosqualodon assenzae)是早中新世的遠洋屬種����。
第二支世係的前附尖傾向縮小 , 主齒尖遠較附齒尖來得大 , 在這支世係中 , 一些屬種如貝氏鯊齒鯨(Squalodon bellunensis)和巴裏鯊齒鯨(Squalodon bariensis)保留了三個後附齒尖 , 這被視作是原始形態 ; 其它屬種有數目眾多的後附齒尖 , 比如卡氏鯊齒鯨(Squalodon catulli)就有多達5至6個後附齒尖 ���。貝氏鯊齒鯨(Squalodon bellunensis)是這世係中原始的代表 , 牠有多達11顆頰牙且上頜後邊的牙齒保留了三個牙根���。較進步的鯊齒鯨屬種比如卡氏鯊齒鯨(Squalodon catulli )有較細小的附尖 , 牙齒也具有數目較多的脊紋���。 巴裏鯊齒鯨(Squalodon bariensis)上頜骨有多達12顆頰齒 , 這是一個較進步的形態 , 但是巴裏鯊齒鯨(Squalodon bariensis)的脊紋數目相對較少 , 附齒尖數目也較多 , 這些特征也被視作是原始形態����。
始鯊齒鯨屬(Eosqualodon)僅隻有兩種包括寬鼻始鯊齒鯨(Eosqualodon latirostris)及朗氏始鯊齒鯨(Eosqualodon langewieschei) , 兩者都有對稱的齒冠 , 齒冠有多達4個前附尖及4個後附尖 , 7至9條長達5 mm的脊紋 , 朗氏始鯊齒鯨(Eosqualodon langewieschei)有多達12顆頰齒 , 但是寬鼻始鯊齒鯨(Eosqualodon latirostris)牙齒數目不明���。始鯊齒鯨屬(Eosqualodon)被認為是原始的鯊齒鯨科成員 , 至於始鯊齒鯨屬(Eosqualodon)那個種是鯊齒鯨屬(Squalodon)的先祖就不得而知 , 也許始鯊齒鯨屬(Eosqualodon)代表第三個世係 , 前附尖及後附尖數目增加���。始鯊齒鯨屬(Eosqualodon)稀疏的脊紋顯示牠們是典型的鯊齒鯨科成員 , 頰齒數目要較一些鯊齒鯨屬種如貝氏鯊齒鯨(Squalodon bellunensis)的多����。
鯊齒鯨屬(Squalodon)在早中新世意大利貝盧諾 (Belluno)一帶的海洋出現 , 能確認早中新世意大利的屬種有兩種 : 巴裏鯊齒鯨(Squalodon bariensis)上下頜骨各有多達58顆牙齒 , 另一屬種貝氏鯊齒鯨(Squalodon bellunensis)上下頜骨則各有多達56顆牙齒 , 至於中新世中期的齊氏鯊齒鯨(Squalodon zitteli)上下頜骨各有多達至少62顆牙齒 ����。中新世的鯊齒鯨屬(Squalodon)有短而套疊的腦殼 , 較大的顳窩及具異型齒 , 中新世的鯊齒鯨屬也有拉長的下頜聯合 ����。鯊齒鯨屬種的頭骨一般長約3呎 , 僅隻是小型的齒鯨類 , 較大的種類能長達16呎���。
凱氏鯊齒鯨(Squalodon kelloggi)在1968年被描述 , 牠的頜骨相當短小 , 頜骨及齒式和其它鯊齒鯨科的有所不同 , 牠被認為是齒鯨亞目位置不定(Odontoceti incertae sedis)����。
鯊齒鯨類在中新世中期至晚期的歐洲及北美開始變得罕見 , 不過在南美早上新世地層也發現懷疑是鯊齒鯨類的化石�����。諾維鋸齒海豚(Prionodelphis rovereti)是一個疑問學名 , 牠的化石被發現於阿根廷早上新世的巴拉那組(Parana formation) , 諾維鋸齒海豚(Prionodelphis rovereti)留下了鯊齒狀的牙齒因此被認為是鯊齒鯨類 , 從牙齒來看牠是具掠食性並可能以魚類或細小的遠洋生物為食 �����。諾維鋸齒海豚(Prionodelphis rovereti)很可能是最後的鯊齒鯨類了���。
鯊齒鯨科(Squalodontidae)曾在地中海及潘諾尼亞海地域生活 , 在羅馬尼亞漸新世地層就曾現過一些被認為是鯊齒鯨屬(Squalodon)的尾椎化石 , 不過這化石是否屬於鯊齒鯨屬(Squalodon)的是很值得懷疑���。確切的化石記錄顯示鯊齒鯨屬(Squalodon)在早中新世阿啟塔階 (23.8-20.5 Ma)及波爾迪階(20.5-18.8 Ma)的地中海及北大西洋曾有過分布 , 這時期地中海 , 印度洋及大西洋都有廣泛的連係����。有一股暖流從中東移向歐洲並把傾向溫暖海洋的無脊椎生物群帶進歐洲 , 潘諾尼亞海中部和地中海之間存有一個海道 , 這個狹窄的海道就是位於現在的北意大利 , 北意大利豐富的生態量也吸引了鯊齒鯨屬在這裏出現 , 比如阿啟塔階北意大利的貝氏鯊齒鯨(Squalodon bellunensis) , 在希臘的阿啟塔階地層也發現了貝氏鯊齒鯨相似種(Squalodon cf. bellunensis)的化石(該化石殘骸保留部分長約378mm)��。在波爾迪階中期至晚期(18.8-16.4 Ma)地中海及印度洋之間的海道封閉 , 但通向大西洋的寬闊海道仍然存在 , 潘諾尼亞海東部失去了和地中海及潘諾尼亞海中部的連接 , 潘諾尼亞海中部迅速退回原來的麵貌 , 在潘諾尼亞海中部的脊椎動物也變得稀少 ���。但是在大西洋東北部及地中海仍是鯊齒鯨類的一片樂土���。
在波爾迪階(Burdigalian)以後的沉積層隻發現少量的鯊齒鯨屬化石 , 牠們都是來自北意大利及德國南部 , 由於地中海和印度之間的聯係已開啟 , 一些來自印太平洋的熱帶動物群落也在地中海及潘諾尼亞海一帶出現 , 地中海及潘諾尼亞海一帶的鯊齒鯨類的沒落主要被認為是和現生齒鯨類出現有關而非環境改變所引起的�����。
在本文為网信彩票論壇精華帖���,由網友兩魏爭雄提供���,僅供大家學習參考��。
編輯��:劉琮瀅
2022-12-03
-

侏羅紀海洋戰鬥俱樂部——上龍
目錄PART I上龍屬—— pliosesaurus macromerus—— pliosesaurus brachyspondylus—— Pliosesaurus brachydeirus—— Pliosesaurus portentificusPART II菱龍屬——Rhomaleosaurus zetlandicus——Rhomaleo
目錄
PART I
上龍屬
—— pliosesaurus macromerus
—— pliosesaurus brachyspondylus
—— Pliosesaurus brachydeirus
—— Pliosesaurus portentificus
PART II
菱龍屬
——Rhomaleosaurus zetlandicus
——Rhomaleosauruscramptoni
巨遊龍屬
——Megalneusaurus rex
PART III
經典���:滑齒龍屬
新發現����:一些尚未定位的巨大上龍科標本
——Liopleurodon ferox
PART IV
巨型上龍的咬合力和生態位
PS���:這次隻包括侏羅紀的上龍���,沒有白堊紀的
世界上最大的上龍是墨西哥上龍型�����,上龍科的阿伯蘭利怪獸��,大概年輕標本是15-18米��,成年標本幾乎可以肯定能有18米����,甚至接近20米��;不過除了阿伯蘭利怪獸之外還有幾種相當龐大的上龍���,目前可達15米的包括Predator-X和巨上龍(pliosesaurus macromerus)
首先介紹上龍屬
上龍屬的分類是相當混亂的����,最早期經常有所謂的巨型上龍的殘片被發現���,一些長頸蛇頸龍型的材料也被認為和大型上龍掛鉤����,接著被命名����,但是大多數最後成了無效種
例如��:Plesiosesarusus giganteus, plesiosesaurus recentior. plesiosesaurus recentior, plesiosesaurus macromerus����,pliosesaurus grandis等
而現在上龍屬尚存四(也許三)個種���,巨型的plesiosesaurus macromerus��,Pliosesaurus brachyspondylus��,體形較小的Pliosesaurus brachydeirus�����,和比較難確定的Pliosesaurus portentificus
它是上龍屬四個種中體形最龐大的���,Kimeridge Clay發現了的巨上龍包括好幾個個體和標本���,其中包括原先被歸納入Stretosaurus����,後來被納入巨上龍種的�����,巨上龍擁有相對在巨型上龍科中最完整的頭後部材料���,最早在1871年被菲利普發現並有效命名���,很長一段時候這個種的大部分大型標本都被稱為Liopleurodon macromerus����,也就是巨大平滑側齒龍���,但是現在卻是已經和平滑測齒龍屬沒有關係了��,最大平滑側齒龍屬也就是殘暴平滑側齒龍���,兩個標本��,一個頭骨中線長度1.26米��,一個據說頭骨(全長����?����,by Tarlo���,不過懷疑和那個中線1.26米的其實是一個個體����,如果是的話就沒有9米了��,最多8米)1.5米���,對應7.5~9����?米的長度���,大概和最小的巨上龍個體差不多大��。
巨上龍的巨大下頜骨
巨上龍分布在英格蘭���,Kimeridge Clay
下麵是最大的巨上龍標本���,一個相當完整的下頜骨
OUM J 10454被說有3米���,實際大小大概2875mm����,不過左邊下頜骨殘缺了一小部分����,修複以後大約略略過3米��,推測體長15米���,或許接近16米�����,但是並不所有的巨上龍都有那麽大的
比如說這頭巨上龍NHM 39362就隻有1700mm��,大概下頜骨總體寬度和長度還不如巴西普鱷和海切爾/葛蘭德恐鱷的最大標本
不過比起其他兩個又大得多了����,Pliosesaurus brachydeirus的材料隻有1085mm���, pliosesaurus brachyspondylus的則是1192mm���,不過後者還有幾個更大的材料��,最大的那個下頜骨大概1734mm
巨上龍和後兩者的主要區別之一就是前頜骨牙齒數量��,後兩者都是10-12顆��,而巨上龍是5-6顆牙���,巨上龍的頜骨比較脆弱��,即使是最大的OUM J 10454前頜骨最寬部分不過280mm���,下頜骨前部比較粗壯�����,最寬牙床部分145mm����,但是後部分隻有75mm寬����。下頜骨24顆牙��,被記載最寬的第5顆大約是75mm牙根寬度����,比最大的霸王龍的牙齒寬度要大35%左右
這次也是巨上龍(雖然標本最初命名不同)
幾個標本的說明���,還包括最大的巨上龍����,巨上龍的普遍長度還是10米多些�����,15米究竟是少數��。而墨西哥阿伯蘭利怪獸基本肯成年個體基本可以達到15米以上
plate 9-4還是最大的巨上龍,其餘的則是兩個當時還稱Stretosaurus. macromerus的化石(J-10441和J35990), 鰭大約是1.85米長���,樓上還可以看到plate 7-3圖上麵標了肱骨�����,股骨和烏喙骨����,Plate 8上麵則是J-10441和J35990經椎骨�����,大約是脊椎柱部分寬度12-14厘米左右����,其中選模的一個頸椎骨高135mm�����,寬138mm����,比最大的恐鱷的中段脊椎骨略大����,卻沒有最大的陸地巨大獸腳亞目的脊椎骨粗����,但比頸椎骨卻要大些��,巨型上龍科成員就最近研究認為可能有軟骨輔助支撐�����,但是這些軟骨保存不下來���。
在另外一個論文裏麵給了J35990還有一些別的材料��,例如這個J35990的肩胛骨部分
巨大的頸椎骨��,大小堪比最大的海切爾恐鱷個體的16節脊椎骨(身體中段)也不過122mmX122mm
上麵還是前段部分的頸椎骨���,後段頸椎骨和脊椎骨更大
巨上龍的肋骨的切麵粗���,長��,而且和脊椎骨融合的位置偏上���,除了脊椎柱的被部分外��,一部分和脊椎骨上部分的Nerual arch接連��。
最後還有一些尾椎骨的數據
脊椎骨長度和體長的比例推測(跟葛蘭德恐鱷對比)的話9.62-11.04米之間
股骨和肱骨的測量數據
現在再推算一下��,這頭J35990的體重和體長����。根據這個圖片作參考
鰭長(肱骨——指末端)長度1839mm��,推測身長11.35米
體寬���:骨盆部分——1490����,前胸部——1510mm
體高���:前胸部——1380mm��,最大部分——1699mm��,
估計大概體重11~12.5噸左右����。
比起最大的巨上龍(15米)個體還是算小的����,但是種群裏麵很肯是平均大小了�����,前麵的那個下頜骨1700mm的可能才9米不到��。
最大的巨上龍大約是28750kg左右����,比最大的霍夫曼滄龍稍微重1噸左右����,不過還有一些計算給出更高的體重���,大概15米����,33589kg(甚至41678kg��,不過這是衛星了)
2003/2004年描述的新上龍屬Pliosesaurus portentificus��,測量尺10厘米��,頭骨長度不會超過最大的巨上龍頭骨的一半��,估計略超三分之一���,保存全長32.2厘米��。也是Kimmeridge Clay Formation����,晚侏羅紀的��。
The mandible measures 332 mm along the midline from the tip of the snout to the posterior of the broken right mandibular ramus.The mandibular symphysis measures 238 mm length, and is 97 mm wide at the posterior
樓上的Pliosesaurus portentificus的標本雖然小���,但是不過是個年輕個體���,很可能是亞成年���,如果SteveEtche私人收藏的下頜骨屬於Pliosesaurus portentificus���,那麽Pliosesaurus portentificus就可能和最大的巨上龍一樣大了���。Pliosesaurus portentificus大概有8顆牙��,正好是巨上龍和Pliosesaurus brachydeirus之間����,brachyspondylus個體變動較大���,大概是9-10顆之間����。
下麵是論文作者的結論���,意思是廢除巨大滑齒龍���,把所有原先屬於巨大滑齒龍的標本劃給巨上龍����;BBC出版《與龍同行》的時候巨大滑齒龍還是有效的����。
現在是 pliosesaurus brachyspondylus��,1840年就發現了一個完整的下頜骨(1192mm)����,以下介紹的標本是1980年發現的����,也是目前最大的標本���,1989一度被認為是滑齒龍屬(天啊��!這麽什麽大型上龍科都被一開始認為是滑齒龍屬��,難道是習慣����?難怪滑齒龍衛星那麽多)���,發現位置是箭頭����。1993年被改為上龍屬��。
最初發現時的位置����,尺子1米��,話說英國人這次挖掘工作做得不這麽樣��,隻收集的主要骨頭��,一些小零碎都被忽略了�����,或者沒有被記錄���。
頸椎骨��,114mm����,大概比前麵的巨上龍J35990的前段頸椎比較大概小了15~20%左右
這個家夥在上龍屬裏麵腦袋算中規中矩���,標本編號BRSMG Cc332大概158厘米�����,其實體積比最大的霸王龍和南巨的頭部還是要小很多的����,海裏麵的也不是都是什麽大怪物�����,雖然和滄龍等比較確實是非常大了��,和18米的霍夫曼滄龍的頭骨差不多��,頭骨可以帶巨大的肌肉群���,估計咬合力不弱���。下頜骨也許比頭骨還要長一些不過可惜缺少了一塊��,它一共有59顆保存下來的頭骨部分的牙齒����,實際推測60顆����,其中左右前頜骨牙10顆��,其餘的是頜骨牙齒
這個家夥在上龍屬裏麵腦袋算中規中矩����,標本編號BRSMG Cc332大概158厘米���,其實體積比最大的霸王龍和南巨的頭部還是要小很多的����,海裏麵的也不是都是什麽大怪物���,雖然和滄龍等比較確實是非常大了�����,和18米的霍夫曼滄龍的頭骨差不多����,頭骨可以帶巨大的肌肉群��,估計咬合力不弱����。下頜骨也許比頭骨還要長一些不過可惜缺少了一塊���,它一共有59顆保存下來的頭骨部分的牙齒��,實際推測60顆����,其中左右前頜骨牙10顆���,其餘的是頜骨牙齒
下頜骨部分����;推測長度173厘米����,根據其他標本複原��,和最大的巴西普魯斯鱷差不多大�����,末端略寬一些
下頜骨左右一共有保存42顆牙齒(牙洞)����,算上前部失去的部分��,下頜骨左右有估計有60~62顆牙齒���,上下總共120~122顆牙齒
前部的大號牙齒的齒冠部分都是5-9厘米之間��,不算特別長��,算牙根�����,最長的233mm���,鋒利����,對成���,穿透型���,但是有一定的切割能力�����,靠近牙尖的部位成刀片���,而底部則是圓形��。
而pliosesaurus brachyspondylus還有另外一種牙齒
較小��,不對稱���,很短�����,帶鉤子����,是靠後麵的牙齒
一共估計有38顆這樣的小型鉤牙����,鉤子不是因為積壓形成���,使原來就這樣的�����,最小的最末端的幾顆也許根本沒有作用���,但是還沒有進化到消失����。
頜骨肌肉組
M. pseudotemporalis 部分的兩塊肌肉負責張嘴和關嘴的動態力量����,M.adductor externus則是合攏以後靜態施加壓力���。
下麵和一個較小的上龍(頭部小����,相對身子和頸部很長)型的菱龍屬Rhomaleosaurus zetlandicus的頭部肌肉比較一下
原創分析一下��:
Rhomaleosaurus zetlandicus的方骨部分傾斜���,M. pseudotemporalis 部分明顯很發達����,而M.adductor externus部分則很弱����,而菱龍屬脖子比較長���,頸部肌肉有力���,這都屬於Warclub式的咬力方式���,也就是說Rhomaleosaurus zetlandicus攻擊獵物時是一開始最大完全張開(看方骨和下頜骨接觸的方式�����,可以確定最大可以張的角度很大)����,然後攻擊����,靠一次性動態咬力關閉嘴巴製服獵物����,M.adductor externus的作用很少
推測
1)對付的是可以一次性擊斃的小型獵物����。直接靠動態咬力
2)這種warclub攻擊方式和異特龍的非常接近
pliosesaurus brachyspondylus則是方骨比較直���,頜骨關節位置可以長大嘴巴���,不能好比Rhomaleosaurus那麽大���,M. pseudotemporalis 可以發動巨大的動態咬力���,而M.adductor externus相當的發達���,咬住獵物以後可以施展巨大的靜態咬力��,前部巨大的穿透型牙齒以動態咬力重創獵物���,稍微彎曲的牙齒控製獵物的掙紮���,接著靜態咬力擠壓��,進一步穿透和殺死獵物���,故此估計pliosesaurus brachyspondylus的獵物比較大����。
文章作者的分析
上龍屬的頭骨前部很平��,頭骨狹窄(用於減少水的阻力)���,頜骨關節強大��,末端咬力很強��,雖然前段咬力未必有多大��,但是對比菱龍屬��,頭骨對掙紮��,尤其是縱向掙紮的獵物的控製力較弱���,也沒有辦法像鱷魚一樣撕下肉����,或許大型上龍類屬於一口吞下大型獵物的掠食者����,它的牙齒也適合對抗大型獵物���。上龍屬是優秀的視覺獵手�����,還有非常發達的水下嗅覺��,小型的溝狀牙齒可以幫助吞咽獵物���,750mm的頭骨寬度足夠吞下一條2-3米的魚龍����。
菱龍屬則有相當強的撕扯能力���,可以肢解體型接近自己的獵物��。
原創評論�����:估計正式因為上龍頜骨對掙紮抵禦較弱����,所以可以才增加咬合力���,迅速增加靜態咬力殺死獵物��,而不是靠被動的增強頭骨抵抗力��,其獵物的絕對平均體形很可能略大於菱龍屬����,通常吃魚龍(在魚龍骨骼裏麵發現了上龍斷裂的牙齒)等較大的獵物���;菱龍屬頭骨較短��,寬����,脖子長�����,咬住的獵物以後可以晃動����,撕扯���,對相對較小的獵物���,菱龍屬也不怕獵物掙紮造成什麽損害��。對付較大的獵物��,warclud的攻擊方式����,一次性打擊獵物以後鬆口����,或者咬下塊肉���,而adductor的作用似乎也不大����,如異特龍就相對很弱���;菱龍屬一般襲擊較小的獵物����,但是偶爾會攻擊和自己差不多大的動物���。
最後分析一下pliosesaurus brachyspondylus的體型���,
BRSMG Cc332�����:158厘米的頭骨��,173厘米的下頜骨���,還有若幹頸椎等���,估計大小時
1)按頸椎骨對比9.45米
2)按下頜骨1:5����,8.7米
3)按頭骨1:6��,9.48米
估計9.2米��,大概體重5噸左右���。
CAMSM 35991���:下頜骨119.2厘米���,估計7.2米左右����,2.6噸����。
上龍亞目的出現在1.98億年���,而菱龍科是侏羅紀早期����,代表的相當特化的上龍����,大型菱龍屬於長頸大頭上龍型���,推測是藍海掠者���,擁有相當好的視覺和水下嗅覺���,縱然曇花一現����,但也為上龍亞目的研究提供了重要的信息下麵向介紹大型菱龍����,Rhomaleosaurus cramptoni, 標本NMING F8785 長度可達7米�����,頭骨中線長度88厘米����,1.78億年前���,1848年出土�����,Whitby, Yorkshire����,英國海岸
頭骨的一些重要區分
(1) rounded dorsomedian foramen situated between the nares(鼻孔中間有開孔)
(2) foramina present on the frontals(頭蓋骨上有開孔)
(3) premaxilla–maxilla sutures run parallel to each other anterior to the nares(前頜骨/頜骨接縫處平行成直線)
(4) palatine excluded from internal naris (choanae),
(5) length and width of premaxillary rostrum subequal,
(6) short mandibular symphysis (length/width=0.60–0.89),
(7) rounded bulb/bump protruding from the medial margin of the retroarticular process
(8) presence of a quadrate foramen.
上麵Henderson等人根據圖片推測Rhomaleosaurus���,3.34米的叉長*[1]個體重482~492kg��,那麽7米個體估測4000~5000kg了���,15米的巨上龍“45短噸”就是這樣推測出來的����,不過嚴重懷疑體重真的有那麽大����。不過其他動物則比較靠普����。
還有就是遊泳的速度明顯不是很快(雖然上麵不是最高速度)����,但是總的來說比魚龍型慢���。
[1]根據下麵這個3.34米的德國標本����:Rhomaleosaurus Victor推測的���,比較草率����,隻能說上龍類在這次研究中不占主體地位��。
頭骨的複原����,缺少了很多材料也對複原造成了困難���,上圖b是馬鐙骨(stapes)��,作者還說應該有一個軟骨的“外馬鐙骨”��,馬鐙骨相當的罕見���,另外著名的標本是白堊紀美國西內海岸的短頸上龍有發現紀錄
Salv部分是外部牙床��,長有完全長成的牙齒���,而正在發育的牙齒則在內部牙床裏麵(Palv)����,當內部牙床的牙齒成長完後就會通過(Can)渠道“遷移”到外部牙床代替外部的牙齒����,前頜骨的牙齒很大���,而上頜骨部分也有一些和前頜骨牙齒大小相近的牙��;
下頜骨相當的完整���,一共有30顆牙齒
下頜骨的寬度和巨型上龍比較相對略顯單薄����,但是相對比例高度缺卻過上龍
前部牙齒的牙根很深���,往後傾斜����,幫助穩固牙齒����,牙齒有些彎曲���,上麵有凸起(比較鋒利)的細痕跡(較小的牙齒則由一些部分沒有這些痕跡)���,目前推測作用是穩固獵物����,幫助穿透還有放血����,牙尖雖然不是很尖�����,但是足夠穿透獵物的皮肉����。
有關於牙齒作用的��,分����,作者的觀點和我前麵24樓的觀點很接近�����,也就是說Rhomaleosaurus的獵物體形跨度很大既可以攻擊大型獵物��,也可以攻擊小型獵物���,巨大的前部牙齒有類似於哺乳動物的犬齒���,深牙根���,彎曲的齒冠幫助控製/撕裂較大的獵物���,將其肢解����,較小的牙齒則負責穩固較小的獵物����,將其吞入喉嚨��,牙齒不尖但很鋒利���,表示可以用來撕扯大型獵物���,或者咬碎小型的甲殼動物���,但是總體上牙齒仍舊屬於切割型的���,前部彎曲的牙齒的最佳實用方式是最大長大是動態攻擊��,或者在大角度咬住獵物發力(相比大型上龍屬則是小角度咬住獵物靜態穿透)��;上下頜骨的牙齒交錯���,而且大小不一���,類似於尼羅鱷等���,而不是食魚鱷
下麵介紹一下北美的巨遊龍屬的Megalneusaurus rex���,老標本19世紀末就發現了���,現在又發現了個新的�����,不果老的標本體形比新的大
北美懷俄明發現��,Sundance Formation��,侏羅紀晚期��,略略在莫爾森階之上�����,新發現的這個標本
最初的情況
Megalneusaurus rex����,君王巨遊龍正模(UW4602)的主要材料包括一個後鰭����,其中長度被推測2米��,發現者Knight曾推測全場2.209米��,股骨長度推測995mm�����,比前麵的巨上龍 OUM J 35990的股骨長3.5厘米���,推測長度至少10米以上���,估計最大可能長度13米���,估測12~13米之間����,這是北美除了阿波蘭利怪獸外幾乎最大的上龍亞目動物了�����。
君王巨遊龍的獵物可能包括了同時期的大眼魚屬的Opthalmosaurus natans���,還有胖胖的蛇頸龍����,Tatenectes laramiensis����,以及Pantosaurus striatus(這個家夥吞下過魚龍胚胎)���,不過當地海洋動物其實最主要的食物還是魷魚����。
平滑側齒龍屬�����,據說這個東西特別“出名”����,幾乎所有的巨型上龍(也許除了predator X) 都被一度歸納入平滑側齒龍屬����,以至於平滑側齒龍極盛一時
以至於出現了這種笑話……隻不過盛極而衰��,最後墨西哥阿波蘭利怪獸和巨上龍都和它沒有什麽關係了��。
平滑側齒龍的頭骨確實威武(同顱骨長度下比克柔龍還給力)����,但是也不能亂放衛星啊
平滑測齒龍到底有多大呢
以下是最近英國發現的一個平滑測齒龍的材料
那麽樓上這頭平滑側齒龍CAMSM J.27424有多大呢����,參考完整的這個英國的平滑側齒龍標本NHM R2680(中線����:前頜骨——方骨長度112厘米)����,那麽大概是中線長123厘米���,全長如果參考俄羅斯的標本[2]133.6厘米����,下頜骨長度144.23厘米��,參考英國本土標本[3]�����,頭骨全長132厘米����,下頜骨141厘米
[2]比例(中線���:全長����:下頜)——(4.86:5.28:5.7)
[3]比例(中線���:全長��:下頜)——(4.05:4.35:4.65)
上麵的推測是根據頭骨����,用小標本(GPIT 1754/2)���,全長4.88米�����,頭骨中線(�����?)長度94厘米,推測CAMSM J.27424長度6.39米����,如果按上龍屬普遍的下頜骨/頭骨1�����:5計算���,則長度最大可能達到7.2米����,實際大小估計在兩者之間���。
可能有人說“平滑側齒龍”沒成年了���,仔細看一下����,上麵說了liopleurodon ferox的“老年個體”NHM R3536頭骨也不過126.5厘米����,下頜骨最大推測148.3厘米���,推測體長6.56~7.6米����,而假設tarlo說的頭骨1.5米的個體存在的話��,長度也不過7.8~8.05米���,體重3~4噸左右��,大概是BBC宣傳的1����:40吧���。
論文的作者估計給BBC給搞怕了��,還特別重申一次說6.4米的平滑側齒龍成年了�����,CAMSM J.27424雖然不是完全融合的那種老年個體����,但是已經沒有軟骨痕跡����,代表骨頭成長期已經基本結束���,是成年個體
最後簡單的說一下斯瓦爾巴特群島上龍和Predator X���,以及一個不知名的大個體
斯瓦爾巴特群島挖出來一個很大的上龍稱“海怪”��,
斯瓦爾巴特群島上龍的頭骨推測2.1米��,下頜骨2.3米����,體長11.5米�����,體長13~14噸���,估計和最大的霸王龍半斤八兩的體重����。比棘龍還小的多
不過如果按鰭長大概接近2.9米算��,根據一些英國方麵的保守推測大概是12.01米����,如果往大了算的話估計是17.3米���,而挪威自己推測15米
1)綜合四個數據大概����:身體部分先2.88*6=17.28米��,乘以0.8��,加上頭骨2.1米(按小頭型算)�����,估計15.9米左右
2)或者四個數據平均13.95米��,大概14-16米之間�����。
體重推測20~35噸之間����。
Erikson認為結構樣子有點像鱷���,但是頭骨實際上幾乎完全和pliosesaurus brachyspondylus差不多���,下頜骨很單薄
Predator X 的碎片
predator X的枕骨推測寬度是霸王龍的2倍���,我們知道霸王龍的枕骨的大小
FNMH PR 2081長9.78厘米���,寬度12.12厘米�����,而ANMH 5027的枕骨大小大概寬12.11厘米和高9.83厘米
那麽Predator X的枕骨估計就是他們的一倍了����,如果直接按霸王龍算��,predator X頭骨長度270~300厘米����,平均285厘米���,枕骨24.2厘米
不過畢竟predator X不是霸王龍���,還是需要用上龍科推測可靠一些��,BRSMG Cc332(pliosesaurus brachyspondylus)的枕骨寬9.286厘米���,而Rhomaleosaurus cramptoni, 標本NMING F8785的枕骨是12.06厘米寬����,也就是說枕骨不是上龍體形大小的唯一標準����,波動是很大����;
Predator X按前者算是頭長4米�����,體長20米的巨型上龍(不可靠)
Prddator X按後者算則是頭長177厘米����,體長14米的菱龍(也不可靠)
不過目前出土的情況媒體的有限消息應該可以確定是頭長3米左右��,或者略不到3米的上龍亞科��,估計體長15米����,體重32.5噸���。
Predator X 15m體
重“45短噸(41000kg)”雖然不是無根據的衛星���,但是可靠程度和埃及棘龍26300-32000kg差不多
“predator X的咬合力視頻上麵介紹是150000N��,媒體上麵說是霸王龍的11倍����,美洲寬吻鱷的13倍”�����,這個肯定是是有錯誤���,前者和最後者是計算錯誤+用詞模糊����,後者是春秋筆法錯誤����。
先說那個150000N的問題
1)視頻裏麵是用來推測的鱷魚猛咬咬力(半動態+靜態���,2組主要肌肉+部分輔助肌肉)�����,單排末端上頜骨牙齒的咬力��,實際雙排半動態咬力要大25%左右����,也就是是187500N���,而2)不是150000N��,動態衝擊咬力就更高了��,但是187500N2)
上龍屬盡管肌肉組合和鱷魚很接近�����,但是頭骨和下頜骨總體結構比鱷魚弱的多���,估計最大咬力(身體衝擊+所有肌肉群)或許勉強187500N���;這不過和最大的巨齒鯊的末端雙排“完全靜態(一組肌肉)”咬力相等��,沒有大的恐怖
霸王龍13400N是一顆牙齒分散咬力(1996年一個案例�����,而且這個不是霸王龍造成的最深的咬痕)���,好幾顆牙一起咬的���,不是像鱷魚那樣把所有的咬力集中在一個接觸點上����,按Rayfield 2004年的分析是估計如果下頜骨不接觸獵物��,單排咬力31000N���,如果下頜骨也算進去45000N��。如果算上雙排消耗��,50000N���,另外霸王龍根據2010年論文上提到的蛇法女妖龍打架那段����,推測能在其他霸王龍下頜骨和頭骨上造成傷口需要29480N的咬力��,奪顆牙齒算入估測至少60000N��,而7噸的平均大小的霸王龍本身的肌肉組合可以提供大概也就70000N的咬力����,算上體重和頸部肌肉組合���,一共可以提供最大咬力125000N���,這個大概等於修改以後的Meer(獵手/獵物咬合力公式)
3)也就是說Predator X的咬力大概隻是7噸的霸王龍大50%��,大概比最大的霸王龍勉強大出20~30%����。加上水的阻力���,而且上龍不可能用上頜骨最後一顆牙咬獵物(前麵說了上龍屬和菱龍屬末端的牙齒又小���,又彎����,還有鉤子��,根本不是用來咬人的)較長的上頜減弱了實際咬力�����,15米的上龍殺傷力估計和最大的霸王龍差不多吧���,這也很正常��,兩者的獵物也差不多大��,甚至是霸王龍的大一些)
至於比鱷魚大13倍�����,估計確實比200-300kg的鱷魚大13倍���,或者比90kg恐爪龍大25倍
言歸正傳�����,上龍一般捕食自己可以吞的下去的獵物�����,或者相對自己體形較小的獵物����,而菱龍科反而可能會去攻擊和自己體形差距較小的��,這個也不算太離譜���,巨上龍一口就可以吞下3米多魚龍了����。而8米的君王巨遊龍的肚子裏麵幾乎都是烏賊的殘骸(石灰質的“烏賊”骨頭)��,最後就是上龍類的牙齒分化基本上很小����,也就是穿透/切碎型�����,不像滄龍科那樣多才多藝��。
再發一套不知名的上龍屬���,但是看它的樣子咬力應該在同顱長的巨上龍個體等之上吧�����,它大概頭骨全長推測2.4米����,體長12米��,也許有~14噸
最後給一個墨西哥怪獸的鰭��,超過3米���,比斯島挪威的還大
大型魚龍那個時候估計是屬於類似於大型上龍那樣吞獵物���,或者撕碎相對自己體形較小的動物����。
早期上龍可能會去襲擊大型魚龍的幼體���,是否會出現群戰還不好說����。大型魚龍估計不會吃上龍成年個體��,吃幼體可以了����,話說其實當時的海洋大型動物也就是這些超大動物��,互相殘殺對方的幼年個體也不是沒可能����,尤其是沒有須鯨類超大型動物����,荔枝魚感覺實在是“孤軍奮戰”又不夠���。君王巨遊龍吃那麽多魷魚/雖然也可能吃魚龍
在本文為网信彩票論壇精華帖��,由網友鴻穹神皇上帝提供����,僅供大家學習參考���。
編輯���:劉琮瀅
2022-12-03
-

滄龍科分類法
滄龍科早期的分類法來源於(Williston, 1897 Russell,1967 Lingham-Soliar, 1992)�����;目前我用的兩套一個是100特征分類法這次用的就是90特征分類法這次發帖先介紹一下前麵的20個特征��,接著剩下的以後再更新滄龍科早期的分類法來源於(Williston, 1897; Russell,1967; Lingham-Soliar, 1992)��;目前我用的兩套一個是100特征分類法
這次用的就是90特征分類法
這次發帖先介紹一下前麵的20個特征��,接著剩下的以後再更新���,不要插樓(如果是說翻譯錯誤等����,集中發表)��,如果有地方覺得翻譯的不對�����,
PS��:Phylogeny And Systematics Of Squamata (Reptilia) Based On Morphology裏麵的500特征分類法太複雜���;懶得去看
這就是前麵的20個特征
不翻譯了��,就是翻譯文字一點用都沒有���,你要查正經點的資料又得轉回英文��,中文什麽都查不到����;翻譯了也是幫倒忙��;到(最前麵的兩個要去龍遊四海(一)裏麵)裏麵可以看到這個表格上所有動物的簡介和翻譯����。
PS����:上麵的巨大滄龍可以當成霍夫曼滄龍的異名
特征1-2
1. Premaxilla predental rostrum absent (0) or present (1).
Premaxillary predental rostrum (if present) short and obtuse (0), distinctly protruding (1) or very large and inflated (2).
1前頜骨最前端牙齒前的突出喙(rostrum)(類似於海諾龍/海王龍屬的撞錘)���;1代表有��,0代表沒有
比如這個就沒斯塔特曼傾齒龍(Prognathodon stadtmani)就是沒有���,牙齒直接出現在最前麵����,索維爾傾齒龍也是沒有這個突出撞錘
2.如果突出喙分存在��,0代表短和不明顯����,1代表比較突出���,2代表相當的突出
特征3-4
3. Premaxilla anteriorly broadly arcuate (0) or narrowly arcuate (1).
 4. Premaxilla internarial bar narrow, distinctly less than half of rostum width (0) or wide, almost as wide as rostrum (1).
3.前頜骨前部��,0代表大弧度擴展���,1代表擴展弧度很小
4 前頜骨內鼻腔部分(internarial bar見下圖)寬度��,0代表狹窄�����,寬度小於突出喙寬度的一半�����,1相當的寬代表超過喙寬度的一半���,或者和突出喙寬度接近���,
霍夫曼滄龍屬於小弧度擴展����,前頜骨內鼻腔部分相當狹窄
佩比海王龍(A)和普氏海王龍(B)則是小弧度擴展(其實差不多所有的滄龍科的可確定屬都是小弧度)
而目前隻有海王龍屬的前頜骨內鼻腔部分(internarial bar)的寬度和喙寬度接近
5. Premaxilla internarial bar base triangular (0) or rectangular (1).
6. Premaxilla internarial bar without (0) or with (1) dorsal keel.
7. Premaxilla internarial bar venter with foramen for fifth cranial (facial) nerve close to (0) or far removed (1) from rostrum.
8. Separate nasals present (0) or absent or fused to other skull element (1).
5.前頜骨比前部分根部���,0代表三角形���,1代表是正方形����,
這個特征不好辨認��,不過說巨大滄龍的是三角型的����,海王龍屬估計是正方形的���。下麵是巨大滄龍前頜骨
6.前頜骨鼻腔背部部分��,0代表沒有‘龍骨’���,1代表有龍骨(原先看錯了���,看成Ventural��,其實是dorsal)����,其實中間凸起的一條細線
7. 前頜骨鼻腔部分的帶有第五頭部神經係統的空腔開孔處����,0代表距離突出喙很近��,
1 距離突出喙很遠
這個特征極為不好辨認���,大部分化石也是(����?)��,很可能壓根沒發現這個部分���,目前隻知道Halisaurus sternbergi這個部位距離���,而普氏海王龍距離相當的遠
8. 鼻腔骨分開��,0代表存在��,1代表和其餘頭骨部分融合�����;這個特征大部分滄龍科和硬椎龍科都是融合的���,但是海王龍屬就沒有融合���,這個可能單看圖也不是很明顯
巨大滄龍就是有龍骨的���,也有一些沒有龍骨的���,但是目前找不到那一部分的俯視圖
這個of Halisaurus sternbergi 就是沒有融合的����,而且前頜骨背上麵龍骨
9. Frontal sides sinusoidal (0) or straight (1).
10. Frontal broad and short (0) or elongate and narrow (1).
11. Frontal not invaded by posterior end of external nares (0) or with distinct narial invasion (1).
12. Frontal midline dorsal keel absent (0), present but low (1) or high, conspicuous and well developed (2).
13. Frontal ala sharp and pointed (0) or more broadly pointed or rounded (1).
14. Frontal olfactory canal not embraced ventrally by descending processes (0) or almost to completely enclosed (1).
15. Frontal overlap on prefrontal indistinct (0) or broad overlap (1).
9.頭蓋骨部分(frontal)��:0代表側麵邊界上呈現三角函數公式形態 ���, 1代表邊界是直線
這個就是三角函數Platecarpus planifrons (YPM 40508).像SIN(X)那樣����,霍夫曼也是���,隻是沒那麽明顯
楚裏傾齒龍的頭蓋骨就是直線
10.頭蓋骨����,0代表寬而且短���,1代表長且狹窄
霍夫曼滄龍����,巨大滄龍��,楚裏傾齒龍的頭蓋骨都是短且寬��,長且狹窄的有近殺手瘤龍和海王龍屬��,以及索維爾傾齒龍���,下圖近殺手瘤龍(左)���,還有普氏海王龍(見以前的圖片)����,不過佩比海王龍卻是寬和短�����,其他傾齒龍也是這樣的����,不一定一個屬裏麵都是一樣的
11.鼻腔股(Nasal)���,0代表沒有突入頭蓋骨����,1代表突頭蓋骨����,霍夫曼滄龍就沒有這麽突入���,但是楚裏傾齒龍就有了����,佩比海王龍非常明顯了
下麵的這幾個也很明顯
12.頭蓋骨表麵中線凸起/‘龍骨’��,0代表沒有��,1代表很矮���,2代表相當的發達��,凸起很高���,這個非常貞觀的了����,很容易就看的出來����,比如霍夫曼就是有凸起的(但不明顯)��,同樣楚裏傾齒龍也有不明顯的凸起
13.頭蓋骨翼�����,0代表形狀鋒利���,尖��, 1代表比較鈍���,呈現較圓 的形態這個特征Halisaurus屬是0��,其他的蒼龍科都是1
Halisaurus sp,可能是指一下這個藍色線條的部位���,其他滄龍這個部位都沒有那麽突出鋒利����。
14.這個特征基本上沒有辦法察看了���,不過指的是olfactory tract(嗅覺管道)�����,下圖的OLt���,0代表沒有向下延伸後被完全封閉�����,1代表幾乎被完全封閉
15.頭蓋骨和前頭蓋骨(prefrontal)接觸部分無法區分為0���,可以區分��,大幅度重疊為1���,這個沒什麽好介紹的��,所有有材料的蒼龍科都是1�����。
16. Frontal posteroventral midline with absence of tabular boss anterior to frontal-parietal suture (0) or presence (1) of tabular boss.
17. Frontoparietal suture with opposing surfaces bearing low interlocking ridges (0) or larger overlapping flanges (1).
18. Frontoparietal suture overlap with oblique medial frontal and parietal ridges contributing to overlap (0) or ridges almost horizontal (1).
19. Lateral sutural flange of frontal posteriorly extended (0) or median sutural flange posteriorly extended (1).
20. Frontal medial invasion of parietal (if present) with short posteriorly extending sutural ridge (0) or long sutural ridge (1).
16.頭蓋骨反麵(ventral)中線在和Parietal接觸部位���,0代表沒有板狀物����,1代表有板狀物,這個沒有一手化石材料的基本上沒法觀察
platecarpus tympaniticus就是沒有板狀物的���,但是海王龍屬的有��,如普氏海王龍��,不過現在也沒有能找到一個海王龍屬頭蓋骨的反麵���。
17. Fronto parietal suture����,頭蓋骨和頂骨接觸縫合線部位��,0代表犬牙交錯的融合���,1較大榫頭組合型融合
這個也沒有什麽好多說的����,基本上都是1���,除了halisaurus這個屬
18. 頭蓋骨和頂骨接觸縫合線部位的正麵切麵��,0融合線代表兩塊骨頭如榫頭一樣交錯���,1代表側視時兩塊骨頭是平行的����;除了海王龍屬的兩個種之外幾乎都是兩塊骨頭交錯的����。這個特征也沒有辦法觀察的
19.這個特征也無法觀測��,省略
20.頭蓋骨突入頂骨(如果有)���,0代表突入部分短����,1代表突入部分長���,���?海王龍屬的普氏海王龍是沒有突入���,tylosaurus nepaeolicus也是沒有突入����,但是這個也沒有法觀察���,除非一手化石數據���。
在本文為网信彩票論壇精華帖����,由網友鴻穹神皇上帝提供���,僅供大家學習參考���。
編輯����:劉琮瀅
2022-12-03
-

肌鱷
拉丁文學名 Sarcosuchus中文名 肌鱷分類 雙孔亞綱 初龍形次亞綱 初龍形類 初龍類 嵌跗類 鱷跗類 鱷形超目 中真鱷類 新鱷類 特提斯鱷類 森林鱷科���?體長 11-12米體重 8噸以上����?時代 早白堊世 Apt拉丁文學名 Sarcosuchus
中文名 肌鱷
分類 雙孔亞綱 初龍形次亞綱 初龍形類 初龍類 嵌跗類 鱷跗類 鱷形超目 中真鱷類 新鱷類 特提斯鱷類 森林鱷科���?
體長 11-12米
體重 8噸以上�����?
時代 早白堊世 Aptian期-Albian期
分布 尼日爾Gadoufaoua的Tenere沙漠的El Rhaz組(El Rhaz Formation)
環境 海岸
食性 肉食
命名 Broin et Taquet, 1966
屬型種 S. imperator 帝王肌鱷
其他有效種 S. hartii 哈氏肌鱷
描述 早在1964年���,法國人拉伯(Albert-Felix de Lapparent)在尼日爾發現了一些巨大的�����,被他戲稱為“鐵軌上的釘子”的牙齒化石���,另外還有一些破碎的骨骼���;這些標本後來被交給了法國古生物學家布萊茵(France de Broin)和塔瑞特(Phillipe Taquet)研究����,他們認為這些化石屬於一種巨大的鱷類���,命名為帝王肌鱷(Sarcosuchus imperator)��,中文有時候簡稱為“帝鱷”����,屬名肌鱷(Sarcosuchus)的意思是“肌肉鱷魚”���。但是因為材料太少��,無法進行有效的複原��,而且法國人又不注意宣傳��,所以這種巨大的動物一直默默無聞���。
但是在2001年�����,這種動物卻一炮走紅��,成了古生物中炙手可熱的“明星”���,這要歸功於美國芝加哥大學的著名古生物學家塞裏諾(Paul Sereno)���。塞裏諾和我們一般印象中整天和死了幾萬年甚至幾億年的動物的骨頭以及不會說話的岩石打交道的��,那種古板����,嚴肅的古生物學家不同���;他是一個善於交際���,喜歡遊走於上流社會����,而且有點喜歡出風頭的家夥����。正是這種性格讓他和大媒體走的很近�����,更是《國家地理》這樣的國際級的媒體結成了同盟��;因此《國家地理》總在第一時間發表塞裏諾的發現����。
不過雖然塞裏諾人前風光���,但是人後吃的苦也不少�����,挖掘帝王肌鱷化石也是一樣���。2000年���,塞裏諾和他手下的一隊人馬闖進被風沙統治的Tenere沙漠��,這個沙漠位於尼日爾的Gadoufaoua����,是把非洲從中間分割成“黑非洲”和“北非”的撒哈拉沙漠的一部分��。雖然��,北非的撒哈拉沙漠一片死寂���,除了偶爾穿越沙漠的穆斯林商隊幾乎沒有人煙��,但是對於塞裏諾這裏是一片“福地”���,他在1996年就曾經光臨過這裏����,並且發現了奇特的三角洲奔龍(Deltadromeus)和比暴龍更加巨大的鯊齒龍(Carcharodontosaurus)的頭骨化石以及幾種鱷類的化石��;這次塞裏諾的一隊人馬也是經過斷水斷糧的折騰��,掉了一身膘才把一些比較完整的帝王肌鱷化石挖掘出來����。
塞裏諾挖掘的的帝王肌鱷化石包括頭骨�����,頸椎和背椎��,背肋����,薦椎和腰帶���,肩胛骨��,頸部到尾部的膜質骨板(Osteoderms)等��;編號為MNN 604—MNN 607����。其中MNN 604為成體��,MNN 605和MNN 606為幼體��,MNN 607為亞成體��。這些標本被送給尼日爾政府���,陳列於尼日爾國家博物館�����。
不過精明的塞裏諾當然不會空手回去���,他找到澳大利亞一家著名的古生物模型公司��,和他們訂下合同��,製作實物大小的帝王肌鱷骨骼模型和複原模型����,根據塞裏諾的推測帝王肌鱷有11到12米長��,體重超過8噸��,所以實物大小的帝王肌鱷模型特別壯觀��;並且聯係了《國家地理》拍攝一部關於帝王肌鱷的記錄片《超級鱷魚(super crocodile)》���,通過這部記錄片�����,人們才真正知道了恐龍時代還有一種這麽巨大的鱷魚���;而且塞裏諾還大膽的推測帝王肌鱷可能捕食恐龍����,這一場景也被《國家地理》用電腦還原���,人們看過以後印象都很深刻��,稱帝王肌鱷是“恐龍殺手”���。
帝王肌鱷屬於新鱷類(Neosuchia)特提斯鱷類(Tethysuchia)下的森林鱷科(Dyrosauridae)��,所謂的森林鱷類都是一些類似長吻鱷的動物����,不過也有人把森林鱷(Dyrosaurus)和肌鱷歸入大頭鱷科(Pholidosauridae)���;帝王肌鱷還有一個不那麽出名的“小兄弟”叫做哈氏肌鱷(Sarcosuchus hartii)���,它們的頭骨略有不同����。
帝王肌鱷的頭骨細長���,但是並不像某些不嚴謹的出版物裏說的那麽誇張��;帝王肌鱷的頭骨長達1.78米��,口鼻部占了大約百分之七十五���,下頜比頭骨略短����。前上頜骨寬大��,有一個圓形的垂瘤����,前上頜骨垂瘤部分比上頜骨寬約百分之二十五����;這個垂瘤部分類似於雄性長吻鱷的Ghara����,不過和長吻鱷不一樣��,垂瘤在所有肌鱷頭骨都有發現���,所以不是第二性征����;所以垂瘤的作用可能是幫助嗅覺���,增加咬合力或者發聲器官���。上頜骨長���,鼻骨特別細長����。隻有一個圓形的外鼻孔���,由前上頜骨的垂瘤圍成��;齶視內鼻孔由狹窄的翼骨分割�����。沒有發現眶前孔或眶前窩(在布萊茵的原始描述中稱發現了眶前孔����,但是MNN 604中沒有發現)���;上顳孔略大於眼眶(Sereno et al., 2001)����。
帝王肌鱷的牙齒圓錐形���,齒冠都具有縱向的棱脊���,橫截麵圓形����,牙齒特別厚實���;雖然這些牙齒並不適合撕裂獵物的批肉��,但是卻很適合固定獵物����,特別是光滑的水生動物��。帝王肌鱷的齒式為前上頜骨齒5���,上頜骨齒30���,齒骨齒31�����,也就是說帝王肌鱷一共有132個牙齒���;在前上頜骨和上頜骨之間有大約十厘米的齒隙�����,齒骨齒的左右間距小於上頜骨齒���。帝王肌鱷屬於“假異齒型動物”���,第三和第四前上頜骨齒(pm3 and pm4)����,第十上頜骨齒(m10)����,第三和第四齒骨齒(d3 and d4)較大���;這些較大的牙齒稱為“假犬齒”�����。
帝王肌鱷的身體覆蓋了很多的膜質骨板�����,所謂膜質骨板也叫“真皮骨”���,是一種外骨骼��,平時我們看到鱷魚背上那些疙瘩一樣的東西就是膜質骨板����。帝王肌鱷最大的膜質骨板位於背部����,有一米多長����,不管是大小還是比例都比現在的鱷魚大得多����。鱷魚的膜質骨板有一種“成長環”��,“成長環”就像樹木的年輪一樣可以記錄生長年齡��;塞裏諾分析了一個大約有最大體型的百分之八十帝王肌鱷的膜質骨板��,這個膜質骨板上大約四十個“成長環”��,也就是說它有大約四十歲��;因此推測帝王肌鱷在五十歲到六十歲之前會一直生長�����。
40歲的帝王肌鱷9米��,30歲的估計6米多���,50歲的10米多���,60歲的11米多���,極限大概是12米���,帝王鱷長得比恐鱷快些��,提前達到最大體型��,也就是50歲的恐鱷9米���,80歲11米����,100歲左右的12米��。
帝王肌鱷MNN係列的大概有6個標本��,GMN係列的至少有1個
Skull of S. imperator (MNN 604). (A) Skull and lower jaws in left lateral view. Cranium in (B) dorsal and (C) ventral views. (D) Anterior end of the jaws in ventral view. Right dentary tooth 4 (E) and 17 (F) in anterior and cross-sectional views. Scale bars: (A) to (C) 50 cm; (D), 10 cm; and (E) and (F), 1 cm. Abbreviations: 1-31, tooth number; a, angular; bo, basioccipital; d, dentary; dep, depression; ec, ectopterygoid; emf, external mandibular fenestra; f, frontal; gr, groove; if, incisive foramen; in, internal nares; j, jugal; l, lacrimal; m, maxilla; n, nasal; oc, occipital condyle; p, parietal; pl, palatine; pm, premaxilla; po, postorbital; prf, prefrontal; pt, pterygoid; q, quadrate; qj, quadratojugal; sa, surangular; sq, squamosal
A) Outline of juvenile (left) and adult (right) snouts of S. (drawn to the same size) (for measurements, see Table 1). (B) Scatter plot and regression lines for total body length versus skull length inGavialis gangeticus (crosses) and Crocodylus porosus (dots) with an estimate of maximum body length for an 87-cm juvenile skull of Sarcosuchus imperator (cross in circle) and the largest recorded adult skull (160 cm) (cross in circle with dashed lines) (18). (C) Strict consensus tree based on three minimum-length trees of 196 steps for 20 crocodyliform taxa based on maximum parsimony analysis of 72 characters (CI 0.52; RI 0.61) (33). Sarcosuchus (bold) andTerminonaris are united by seven unambiguous synapomorphies (node 2); another six unambiguous synapomorphies unite these taxa,Pholidosaurus, and Dyrosaurus (node 1) (characters and data matrix available at Science Online) (29). Dots at nodes for Crocodyliformes and Metasuchia signify node-based definitions of node-stem triplets (30). (D) Skull of Terminonaris robusta (98 cm length) in dorsal view (right), with the end of the snout shown in right lateral (top left) and ventral (bottom left) views. Scale bars in (D), 10 cm.
Skeletal reconstruction of adult S. imperator showing (A) preserved bones (right side) and scutes (left side) (length 11.65 m, or 38 feet, 3 inches). Individual bones: (B) mid cervical vertebra in left lateral and anterior views (juvenile MNN 605), (C) posterior dorsal vertebra in lateral and anterior views (subadult MNN 606), (D) mid cervical scute in dorsal view (subadult MNN 607), (E) trunk scute in dorsal view (MNN 605), (F) caudal scute in dorsal view (MNN 607), and (G) pelvic recon- struction (juvenile MNN 606, subadult MNN 607). Scale bars in (B) to (G), 10 cm; human silhouette is 1.68 m (5 feet, 10 inches) tall. Abbreviations: acet, acetabulum; ant, antitrochanter; apr, anterolateral process; di, diapophysis; il, ilium; is, ischium; k, keel; ms, median suture; nc, neural canal; ns, neural spine; om, overlap margin; pa, parapophysis; pop, postacetabular process; poz, postzygapophysis; prp, preacetabular process; prz, prezygapophysis; pu, pubis; tp, transverse process.
最大的帝王肌鱷MNN604隻有頭骨�����,其他都是從小個體拚起來的��,MNN 605是確定幼年����,其餘都是沒有完全長成�����,或者“亞成年”
可以看出小個體頭部比例很大���,較大個體比例較小��;帝王肌鱷按恒河鱷裏麵超過3米的個體的頭部身體比例推測大概是10-11米���,按最大的一個推測是10.56米�����,而按灣鱷超過3米的個體的比例推測11~12.5米����,最大的一個推測是12.5米��;
按下麵的公式則是
帝王鱷正模全長度是160厘米��,還有一個170厘米(HL~160厘米)����,那麽全長170厘米的按灣鱷推測12.15米��,按恒河鱷11.15米��,
體重7960kg則是按12.15米灣鱷推測的���,所以不是超過8噸�����,是最大8噸��。
MNN G 102-2大概7.25~9.107米���,比它大33%的帝王肌鱷大約是9.64~12.13米����,分別和灣鱷的上限���,恒河鱷的下限吻合
最大的帝王鱷也不是都是8噸的推測��,帝王肌鱷MNN G102-2按照野生密河鱷(*代表是按完全按照野生個體估算的)����,估算的長度9.1米(見樓上表格���,攜帶*)����,體重2410kg���,比它大33%的最大帝王肌鱷(MNN 604)大概按股骨周長增長指數(2.125~2.85)推測大概是4417~5432kg���,這個大概和FNMH PR 2081霸王龍5543~6197kg��,南巨5390~5653kg或者棘龍8845~10181kg差不多�����,如果帝王鱷8噸����,大概對應FNMH PR 2081 10噸��,棘龍20噸��。
在本文為网信彩票論壇精華帖��,由網友三千院凪提供��,僅供大家學習參考��。
編輯���:劉琮瀅
2022-12-03
-

旭日東升的逆戟王朝
逆戟鯨屬(Orcinus)是一個具有悠久曆史的真海豚科家族 , 在意大利及日本上新世地層都發現過許多逆戟鯨屬(Orcinus)的牙齒 , 在新西蘭早更新世地層也發現過逆戟鯨相似屬(cf Orcinus)的牙齒化石 ����。在意大利早
逆戟鯨屬(Orcinus)是一個具有悠久曆史的真海豚科家族 , 在意大利及日本上新世地層都發現過許多逆戟鯨屬(Orcinus)的牙齒 , 在新西蘭早更新世地層也發現過逆戟鯨相似屬(cf. Orcinus)的牙齒化石 ��。在意大利早上新世地層發現的西托克逆戟鯨(Orcinus citoniensis)有良好的頭骨保存 , 每邊顎骨保存的牙齒有14顆 , 西托克逆戟鯨(Orcinus citoniensis)體長一般不足4米 , 牠的形態介乎在真海豚科及現存的逆戟鯨之間����。除了西托克逆戟鯨(Orcinus citoniensis) , 目前對上新世至更新世的逆戟鯨屬(Orcinus)的認識非常稀少 , 這5百萬年的時間肯定是逆戟鯨屬(Orcinus)的黃金時期 , 可是上新世至更新世地層出土的化石太稀少 , 但是近來對幾個地域的逆戟鯨個體進行線粒體的基因組分析顯示逆戟鯨在更新世時期有過分化 , 現在的逆戟鯨屬可能有超過4種甚至更多 , 本文會先介紹北美過客型 , 居留型 , 遠洋型及其它地域的逆戟鯨習性和食性 , 然後再簡略介紹牠們彼此的關係 , 在本文中不會對什麽何年何月何日的冗長捕食記錄及牠們的基本數據比如什麽平均體長和妊娠期等等作詳細的介紹���。西托克逆戟鯨(Orcinus citoniensis)
東北太平洋的逆戟鯨除了過客型 , 居留型及遠洋型以外 , 還有其它身份不明的逆戟鯨比如LA小群���。在這裏先介紹一下過客型 , 居留型及遠洋型���。
過客型逆戟鯨有較尖的背鰭 , 牠們的鞍狀紋沒有分叉 , 過客型小群一般由1至7頭個體組成 , 平均就隻有3至4頭 ���。過客型逆戟鯨的移動模式難以估計 , 但是牠們傾向在海岸線逗留 ���。過客型逆戟鯨的小群包括成年母鯨及牠的後代(後代一般為一至兩頭) , 成員之間彼此有持久的關係, 雄鯨和牠們的母親維持緊密的關係並持續至成年期���。但是一些雄鯨會離開牠們的母係群並成為流浪的雄鯨 , 這些流浪的雄鯨不會和其它逆戟鯨形成長久的群體 , 僅隻是偶爾加入其它雌鯨群體進行繁殖 , 這些流浪的雄鯨更不會和其它雄鯨存有什麽聯係���。雌鯨就算離開牠們的母係群 , 也會比雄鯨更具群居性 ���。過客型逆戟鯨母係群會和其它過客型逆戟鯨的母係群作短暫性的聯係 , 牠們彼此不會形成長遠的關係��。
在偵察獵物時過客型逆戟鯨會發出極低頻率的聲波 , 由於逆戟鯨有良好的雙眼視覺 , 牠們的舌頭對化學物質也相當敏感 , 這也提供了偵測獵物位置等訊息 , 過客型逆戟鯨在接近獵物時主要依賴視覺及聽覺 ����。過客型逆戟鯨主要以海生哺乳類為食 , 牠們的食物主要是鰭足類及鯨類, 什至連北美水獺(Lontra canadensis)及鳥類也偶爾是過客型逆戟鯨的餐單內����。
研究顯示太平洋東北部存在了三個基因上有差異的群落 , 分別是1)西岸過客 , 分布於加利褔亞南部至阿拉斯加東南部 ; 2)阿拉斯加海灣過客及3) AT1群落 , 分布在阿拉斯加灣北部的奇奈峽灣及威廉王子灣(prince william sound), 目前AT1群落頻臨滅絕���。基因上的研究顯示這三個過客型逆戟鯨的群落隻存有極少或者根本沒有什麽基因交流����。在太平洋西北部的日本海岸以外估計有多達 1200頭逆戟鯨 , 西北太平洋的逆戟鯨可分作過客型及居留型 , 該區域的過客型逆戟鯨也有捕食北海狗(Callorhinus ursinus)及海象(Odobenus rosmarus) , 攻擊小須鯨(Balaenoptera acutorostrata)及座頭鯨(Megaptera novaeangliae)的記錄�����。在日本海岸記錄了最大的雄鯨長達9.8米 ��。
居留型逆戟鯨有鉤狀的背鰭 , 背鰭尖呈圓狀 , 居留型逆戟鯨小群由10頭至60頭個體組成����。居留型逆戟鯨主要以硬骨魚類為食 , 捕食時會發出相當頻繁的卡嗒聲 , 這些聲納能對魚類構成傷害����。居留型逆戟鯨可分作五個群落 , 包括北方居留型 , 南方居留型 , 南阿拉斯加居留型 , 西阿拉斯加居留型及西北太平洋居留型���。
南方居留型逆戟鯨有三個小群包括J小群 , K小群及 L小群 , J小群及K小群大約有多達20頭成員 , L小群更多達59頭成員 , 占去整個南方居留型種群總數的一半��。在春季 , 夏季及秋季南方居留型逆戟鯨小群在普捷灣(Puget Sound) 內陸水道 , 喬治亞海峽(太平洋海灣)及南側隔海峽(STRAIT OF JUAN DE FUCA)出沒 , 這些小群定期在華盛頓及溫哥華島海岸出沒 , 並且罕有地南移至加利褔亞中部及北移至夏洛特皇後群島(Queen Charlotte Islands)���。至於冬季時南方居留型逆戟鯨的種群分布及移動所知什少�����。盡管南方居留型及北方居留型在北麵的溫哥華島及夏洛特皇後群島(Queen Charlotte Islands)分布上有所重疊 , 但是南方居留型及北方居留型的小群並沒有被發現具任何基因上的交往 , 基因上的分析顯示南方居留型及北方居留型有著生殖隔離���。
南方居留型逆戟鯨雖然主要以鮭魚及其它魚類為食 , 但要較其它居留型逆戟鯨凶猛 , J小群更有襲擊殺死港灣鼠海豚的記錄 ����。2005年的報導指出在聖璜島(San Juan Island)發現過13頭港海豹 (Phoca vitulina)的屍體被認為可能和南方居留型逆戟鯨有關 , 近來在哥倫比亞省納奈莫市更有目擊多達14頭南方居留型逆戟鯨攻擊及追逐3頭過客型逆戟鯨���。
北方居留型逆戟鯨群落一共有16個小群 , 牠們生活於溫哥華島(Vancouver Island)中部至阿拉斯加東南部 , 偶然也會在南方的德福卡海峽 , 聖胡安島嶼及Gray’s Harbor出沒���。從六月至十月 , 許多北方居留型逆戟鯨小群會聚集在溫哥華島東北麵的約翰斯通海峽(Johnstone Strait)及夏洛特皇後灣(Queen Charlotte Sound) , 但是對北方居留型逆戟鯨在其它月份的移動及分布所知什少���。在阿拉斯加東南部 , 北方居留型會和南阿拉斯加居留型有所接觸但兩個群落之間卻沒有或者極少的基因交流�����。
南阿拉斯加居留型逆戟鯨分布在阿拉斯加東南部及阿拉斯加海灣[包括威廉王子灣(Prince William Sound)及科迪亞克島(Kodiak Island)] , 南阿拉斯加居留型一共有至少15個小群在兩個區域被確認��。威廉王子灣(Prince William Sound)的居留逆戟鯨並沒有出沒於阿拉斯加東南部 , 但是在科迪亞克島(Kodiak Island)外和其它未知的居留逆戟鯨小群混集����。
至於西阿拉斯加居留型逆戟鯨更所知什少 , 牠們的分布僅局限在白令海一帶的海岸及遠洋���。曾在西阿拉斯加拍攝得多達241頭逆戟鯨 , 但是牠們的小群數目卻是未知的����。估計有多達400頭逆戟鯨分布在白令海東南部 , 但是沒法確定白令海東南部到底存有多少頭居留型逆戟鯨�����。
西北太平洋居留型分布在華盛頓 , 不列顛哥倫比亞省及阿拉斯加的海岸 , 這也是三文魚的分布����。西北太平洋居留型的分布可能一直向西延至俄羅斯和日本的海岸線��。
遠洋型逆戟鯨形態和居留型逆戟鯨要較相近 , 遠洋型逆戟鯨的鰭尖較圓狀 , 鞍狀紋大小和居留型逆戟鯨的相近 , 但是鞍狀紋較不明顯 , 而且鞍狀紋的變異較過客型的大 ��。遠洋型逆戟鯨規模要遠較過客型及居留型的大 , 一群遠洋型逆戟鯨的數目能多達75至100頭個體 , 盡管過客,居留及遠洋逆戟鯨三者的分布有所重疊 , 但遠洋型逆戟鯨和過客型或居留型逆戟鯨彼此沒有混集���。遠洋型逆戟鯨也是三者中體型最細小的 , 有記錄的遠洋型雄鯨全長約6.2至6.5米 , 雌鯨全長約5.5至5.7米�����。遠洋型逆戟鯨偏好以硬骨魚及鯊類為食 , 牠們的食物包括虹鱒屬(Oncorhynchus sp.) , 杜父魚屬(Cottus sp.) , 狹鱗庸鰈(Hippoglossus stenolepis) , 月魚(Lampris guttatu)及真鯊科比如大青鯊(Prionace glauca)��。遠洋逆戟鯨也被目擊和加州海獅(Zalophus californianus), 灰鯨(Eschrichtius robustus) , 長須鯨(Balaenoptera physalus) 及短吻真海豚(Delphinus delphis)同遊 , 但並沒有任何證據顯示是存有捕食關係 , 什至還有目擊記錄顯示瑞氏海豚(Grampus griseus)向一對遠洋逆戟鯨不斷進行騷擾 ���。
在夏威夷的逆戟鯨基因則和阿拉斯加海灣的過客型逆戟鯨較接近 ,估計有250頭逆戟鯨在夏威夷一帶的海洋生活��。
有關加利褔亞以外的逆戟鯨習性及食性所知什少 , 著名的那個法拉隆島殺死大白鯊的LA小群到底和其它地域的逆戟鯨比如過客,居留及遠洋的關係怎樣一無所知 , 這個LA小群看似以海獅及鯊類為食���。在墨西哥海灣的逆戟鯨曾有捕食熱帶斑點原海豚(Stenella attenuata)的記錄����。在巴哈馬以外的大西洋有目擊過逆戟鯨捕食大西洋點斑原海豚(Stenella frontalis) , 弗氏海豚(Lagenodelphis hosei)及侏抹香鯨(Kogia sima)的記錄��。
西北大西洋的逆戟鯨主要以琴海豹(Pagophilus groenlandica) , 白喙斑紋海豚(Lagenorhynchus albirostris) , 小須鯨(Balaenoptera acutorostrata) , 白鯨(Delphinapterus leucas), 座頭鯨(Megaptera novaeangliae) , 刀嘴海雀(Alca torda) , 藍鰭金槍魚(Thunnus thynnus)及大西洋鯡(Clupea harengus)為食�����。
在加拿大東部的北極地域的逆戟鯨以弓頭鯨(Balaena mysticetus) , 白鯨(Delphinapterus leucas) , 一角鯨(Monodon monoceros) , 長肢領航鯨(Globicephala melas) , 長須鯨(Balaenoptera physalus) , 小須鯨(Balaenoptera acutorostrata) , 座頭鯨(Megaptera novaeangliae)及海豹為食���。
北大西洋的逆戟鯨可分作1型(Type 1)及2型(Type 2) , 過去認為牠們和東北太平洋逆戟鯨及南極逆戟鯨彼此獨立分化 , 但是近來的基因分析顯示大西洋2型逆戟鯨和南極的逆戟鯨關係更密切 , 北大西洋1型及北大西洋2型是屬兩個不同的種群��。
北大西洋1型由七個單倍體型(單倍體型為29–35)組成 , 在冰島及挪威牠們以鯡魚為食 , 在北海的北大西洋1型則以鯖魚為食���。北大西洋1型有較平行的眼斑 , 眼斑前端正是在噴水孔前方 , 北大西洋1型雄鯨的體長平均數為6.3米 , 所捕獲的最大個體長約6.5米�����。
北大西洋2型分布在蘇格蘭至法羅群島(介乎挪威海和北大西洋中間) , 北大西洋2型有呈三角型的眼斑 , 眼斑前端的位置是在噴水孔後方 , 在蘇格蘭西岸及亞速爾群島的小型群落發現了色素圖樣 ����。北大西洋2型雄鯨的體長平均數為8.16米 , 最大的個體可達8.5米(據說在東北大西洋曾捕獲9米的個體)���。北大西洋2型由兩個單倍體型(單倍體型為27及28)組成 , 和北大西洋1型不同 , 北大西洋2型主要以其它鯨類為食的 , 曾在牠們的胃部發現了小須鯨(Balaenoptera acutorostrata)的鯨須 ���。在墨西哥灣襲擊及捕食抹香鯨的單倍體型37則被認為是屬北大西洋2型���。
挪威北部的逆戟鯨以大西洋鯡(Clupea harengus)為食時 , 一群逆戟鯨會把鯡魚群趕往水表層 , 鯨群會在魚群四周或者下方遊動 , 定期地用尾巴拍打魚群才進食�����。挪威北部的逆戟鯨群偏好在早上於淺海尋找鯡魚群 , 牠們甚至會跟隨漁船並進食墮進魚網的鯡魚���。
冰島的逆戟鯨發展出另一種捕獵策略 , 牠們會發出聲波把鯡魚群趕在一起, 進食前再以尾巴拍打鯡魚群���。
在蘇格蘭北部及挪威之間的水域逆戟鯨更被目擊在漁船附近追蹤大西洋鯖魚(Scomber scombrus)和鯡魚群 , 在白令海更有目擊逆戟鯨小群尾隨漁船多達31天且長達1,600公厘���。
在直布羅陀海峽(Strait of Gibraltar)的逆戟鯨群以追捕遷徙的藍鰭金槍魚(Thunnus thynnus)為食 , 牠們以每秒3.7米的時速持續追逐藍鰭金槍魚(Thunnus thynnus)達30分鍾 , 直布羅陀海峽的逆戟鯨主要以體長小於1.5米的藍鰭金槍魚(Thunnus thynnus)為食 , 至於體型較大的藍鰭金槍魚(Thunnus thynnus)對逆戟鯨來說是很難追捕的獵物 ����。
在新西蘭不同的逆戟鯨亞群落都存在不同的食性 , 整體而言牠們的小群由2頭至22頭個體組成 , 新西蘭逆戟鯨小群大小和食物類型有很大關連 , 以鰩類為食的小群大小一般由3至18頭個體組成 , 以鯨類為食的小群大小則由3至6頭個體組成 , 以鯊類為食的小群大小則由5至8頭個體組成 ����。新西蘭逆戟鯨大概和東北太平洋的逆戟鯨相似 , 都是比較偏食的 , 一些小群專門以鰩類為食 , 另一些則專門以鯨類為食 , 不過也許牠們可能也是機會主義者����。
牠們的食性大致包括軟骨魚 , 硬骨魚 , 鯨類 , 鳥類及頭足類(主要是章魚)��。新西蘭逆戟鯨曾有捕食翅鯊(Galeorhinus galeus) , 大青鯊(Prionace glauca)及短肢灰鯖鯊(Isurus oxyrinchus)的記錄 , 牠們的食物還包括鰩類 , 什至曾有捕食紐西蘭電鰩(Torpedo fairchildi)的記錄 �����。新西蘭逆戟鯨捕食的硬骨魚主要包括黃鰭金槍魚(Thunnus albacares) 及南極櫛鯧(Hyperoglyphe antarchia) , 曾有目擊記錄顯示逆戟鯨曾捕食翻車魨(Mola mola)�����。新西蘭的逆戟鯨捕食的哺乳類包括暗色斑紋海豚(Lagenorhynchus obscurus) , 曾有過襲擊短吻真海豚(Delphinus delphis) , 南方露脊鯨(Eubalaena australis)及座頭鯨(Megaptera novaeangliae)的記錄����。令人奇怪的是鰭足類不是在新西蘭逆戟鯨的餐單內 , 對新西蘭的鰭足類來說 , 大白鯊(Carcharodon carcharias)是主要的威脅����。至於新西蘭的逆戟鯨捕食的鳥類主要是小藍企鵝(Eudyptula minor), 至於黑背鷗(larus dominicanus)及新西蘭紅嘴鷗(Larus scopulinus)並不是新西蘭逆戟鯨的食源之一 , 僅隻目擊過新西蘭遠洋的逆戟鯨偶然捕食黑背鷗(larus dominicanus)及新西蘭紅嘴鷗(Larus scopulinus)而已����。
南極南部正是逆戟鯨的集中地 , 當地有三類逆戟鯨 , A型逆戟鯨主要以鯨類為食 , B型逆戟鯨主要以鰭足類為食 , B型逆戟鯨可能也會以企鵝作為食物 , C型逆戟鯨以硬骨魚為食�����。
根據1983年Berzin 及Vladimirov的觀察, 南極遠洋型的逆戟鯨大致上是A型 ,小群數量估計都隻是10-15頭����。 B型及C型主要活動於浮冰附近 , 根據Berzin 及Vladimirov的觀察浮冰附近有多達150-200頭逆戟鯨,但他們沒把這150-200頭逆戟鯨區別那些是B型及C型 , 因此B型及C型種群數量是不太清楚的,但好像比較傾向C型 ; 至於1981年Ivashin觀察浮冰附近已有多達 150-300頭逆戟鯨,但也沒指出B型及C型的數量����。
其實在南極還發現了第四類逆戟鯨 : D型逆戟鯨 , D型逆戟鯨最初在1955年的新西蘭被拍攝 , 在2004年以後有六次目擊D型逆戟鯨的記錄���。D型逆戟鯨有極細小的白色眼斑 , 牠分布在緯度40°S 及 60°S之間亞南極水域 , 牠們的頭部更呈球狀並類似領航鯨 , D型雄鯨有極狹窄的背鰭並向後傾斜 ���。目前對D型逆戟鯨的食性還是不明 , 但是有照片顯示D型逆戟鯨會掠奪漁船捕獲的小鱗犬牙南極魚(Dissostichus eleginoides)���。其實南極海洋應該還有第五類逆戟鯨 , 下文將會再描述���。
南半球的逆戟鯨比較偏向進食須鯨的唇及舌頭部分,北半球的逆戟鯨不但進食須鯨的舌及唇部分,連咽喉位置也是北半球的逆戟鯨偏好的進食部分���。據說在南半球的兩頭雄性逆戟鯨胃部也發現過其它逆戟鯨的殘骸 ��。
有記錄顯示南非的逆戟鯨以硬骨魚類及長吻真海豚(Delphinus capensis)為食 ����。
於大西洋西南部的逆戟鯨則會同時以魚類及海生哺乳類為食 , 在巴西南部以外的水域及世界許多地區的逆戟鯨都懂得捕食困在魚網的魚類 , 在南大洋逆戟鯨出沒時 , 小鱗犬牙南極魚(Dissostichus eleginoides) 漁獲減少超過50%����。在委內瑞拉東北部的海岸也目擊過逆戟鯨捕食太平洋橄欖鱗海龜(Lepidochelys olivacea)及棱皮龜(dermochelys coriacea)的記錄�����。
在巴西São Paulo北岸的淺海有過逆戟鯨捕食雙吻前口蝠鱝(Manta birostris)的記錄 , 巴西的東南部及南部也有逆戟鯨捕食劍魚科及金槍魚的記錄��。
阿根廷瓦爾德斯半島以外的逆戟鯨什至懂得”擱淺戰術”捕捉海獅 ; 在阿根廷丘布特省海岸逆戟鯨有攻擊捕殺七鰓鯊(Notorhynchus cepedianus)的記錄 ��。
秘魯的逆戟鯨偏好以海生哺乳類為食 , 曾有目擊記錄顯示有6頭逆戟鯨曾追逐一群有15頭個體的領航鯨小群 ,另一個記錄是8頭逆戟鯨追逐多達200頭寬吻海豚(Tursiops truncatus) , 牠們殺死了其中一頭寬吻海豚(Tursiops truncatus)並多次把海豚撞出水麵 ����。在當地的抹香鯨身上發現的傷痕也被認為是秘魯的逆戟鯨所做成的����。秘魯的逆戟鯨也有捕殺南美海狗(Arctocephalus australis)幼體的記錄 ; 也有南美海獅(Otaria flavescens)被發現和逆戟鯨相距800米 , 但是卻沒顯示南美海獅(Otaria flavescens)有逃避的跡象���。
加拉帕戈斯群島的逆戟鯨有捕食抹香鯨(Physeter macrocephalus)及布氏鯨(Balaenoptera edeni)的記錄 , 牠們的食物包括硬骨魚類 , 軟骨魚類 , 海獅科及鯨類 , 被捕食的軟骨魚類包括加拉帕戈真鯊(Carcharhinus galapagensis) , 雙髻鯊屬種(Sphyrna spp.) , 雙吻前口蝠鱝 (Manta birostris) , 太平洋前口蝠鱝( Manta hamiltoni) , 魟屬種(Dasyatis sp.)及鱝屬等等���。
曾拍攝未成熟的逆戟鯨會碰撞雙吻前口蝠鱝 (Manta birostris)並把獵物向下拖至深海 , 逆戟鯨會把蝠鱝殺死並水平地遊至另一頭逆戟鯨 , 也有一個水下短片記錄小群中成年的逆戟鯨和未成熟的逆戟鯨共同撕裂懷疑是鱝科的獵物���。在加拉帕戈斯群島一帶的海洋經常都可目睹逆戟鯨獵殺雙吻前口蝠鱝 (Manta birostris) , 此外芒基蝠鱝(Mobula munkiana)及納氏鷂鱝(Aetobatus narinari)也是加拉帕戈斯群島逆戟鯨的餐單中 , 這些大型鱝科 (Myliobatidae)屬種看似是加拉帕戈斯群島逆戟鯨的主要食源 , 但是加拉帕戈斯群島逆戟鯨也多次被目睹和加拉帕戈海獅(Zalophus wollebaeki)的領域相當接近 , 這暗示逆戟鯨應該會捕食加拉帕戈海獅(Zalophus wollebaeki) , 並顯示牠們是機會主義者���。相對鯨類及海獅科這些高潛能的獵物 , 大型鱝科遊速較低 , 逆戟鯨要捕食大型鱝科相對要較容易且不用付出大多體力 , 此外捕食這些大型鱝科也是成鯨教導未成熟的個體捕獵技巧的理想對象��。
印度洋的逆戟鯨數量不明 , 在印度洋的逆戟鯨有捕食金槍魚及鯊類的報導���。在克羅澤群島(Crozet Islands)一帶的印度洋至少存有兩類逆戟鯨 , 當地有一類逆戟鯨外型和南極的A型逆戟鯨相似 , 牠們的食性相當廣泛 , 曾被目擊以小須鯨 , 南象海豹 , 企鵝及魚類為食 ; 那裏的A小群中的雌鯨成員A2 及 A3長度分別為6.1米及5.9米 , 這和挪威的逆戟鯨雌鯨平均大小差不多 , 挪威的雌鯨體長平均數為5.8米 , 最大的為7米 ����。冰島發現的雌鯨最大則為5.4米���。
生活在克羅澤群島(Crozet Islands)及凱爾蓋朗群島一帶印度洋的另一類逆戟鯨則被目擊以小鱗犬牙南極魚(Dissostichus eleginoides) 為食 ����。上述的南極 D 型逆戟鯨則有14個在克羅澤群島(Crozet Islands)被目擊的記錄 , 但是這些記錄都是發生在遠洋 , 那裏的小鱗犬牙南極魚(Dissostichus eleginoides)正被認為是南極 D 型逆戟鯨的食物���。
澳洲的逆戟鯨食性相當廣泛 , 食物包括硬骨魚類 , 鯊類 , 企鵝 , 儒艮 , 鰭足類及鯨類 , 什至連抹香鯨也曾受澳洲逆戟鯨襲擊進食���。
幾乎所有南極的逆戟鯨類型是一個單源類群 , 這些南極的逆戟鯨類群和生活在西北大西洋墨西哥海灣的逆戟鯨種群形成單源的類群 , 牠們和東北太平洋的過客型逆戟鯨關係並不密切 , 相反牠們和東北大西洋及東北太平洋的其它逆戟鯨關係非常密切���。南極B型及C型逆戟鯨是有共同先祖 , 從線粒體的基因組的分析研究顯示南極B型及C型逆戟鯨僅隻有三個差異 , 南極A型逆戟鯨和B型及C型逆戟鯨卻有著24至25個差異 ���。值得一說的是南極其中一個逆戟鯨種群AntA2和其它南極逆戟鯨絲毫沒有任何關係 , AntA2是一個類似過客型的獨立種群 , AntA2和東北太平洋的過客型逆戟鯨關係要非常密切 , 牠是南極第五個逆戟鯨類群��。
過客型逆戟鯨種群由牠們的共同先祖分化估計約有19萬年 , 其它逆戟鯨類群分化估計約5.9萬年至11.7萬年 , 南極的逆戟鯨類群和墨西哥海灣的逆戟鯨種群由牠們的共同先祖分化估計約33萬年之久���。
遠洋型逆戟鯨包括東北太平洋的遠洋型類群外 , 還包括墨西哥西部及克利珀頓島(位於東太平洋墨西哥海域的荒島)的逆戟鯨種群(這兩個種群過去沒被認為是遠洋型逆戟鯨的一部分)及西北大西洋紐芬蘭的一個逆戟鯨種群 , 這顯示東北太平洋的遠洋型及居留型都源自北大西洋 , 或者是在氣候回暖時從東北太平洋穿過西北海道返回西北大西洋��。
ENA 1-2這分化枝主要有兩個不同的單元型(haplotypes) , 主要分布在英國及冰島一帶的海洋 , 牠們曾被列入北大西洋1型(North Atlantic Type 1)����。ENA type 1主要以魚類為食 , 但牠們被認為同時以魚類及哺乳類為食 ���。ENA 1-2兩個不同的單元型的分化時間大致和東北太平洋的居留型及遠洋型分化時間相同 ���。
ENA Type 1這分化枝包括冰島 , 挪威及直布羅陀海峽的逆戟鯨及來自新西蘭其中一個群落 , ENA Type 1是近來根據食性和型態而被描述的 ��。從分析研究顯示ENA Type 1和東北太平洋的居留型及遠洋型關係要較密切 , 東北大西洋及東北太平洋兩地的逆戟鯨有著相似的單元型分化 , 但是沒法知道兩地群落的共同先祖到底是源自北大西洋或者是北太平洋 , 牠們可能興起於低緯度區域 , 但這些還需要更多的研究支持���。
ENA Type2僅隻有一個樣本被抽取研究 , 從分析研究顯示ENA和其它的東北大西洋逆戟鯨群落有很大差異 , 但是牠們之間的關係還需要更多的分析研究����。
根據研究分析顯示北太平洋的過客型逆戟鯨應該被認為是獨立種 , 北太平洋的過客型逆戟鯨從其它逆戟鯨分化出來已有70萬年之久 ; 南極B型及C型也被認為是獨立種 , 牠們的分化時間已多達15萬年之久 , 南極B型及C型兩者和南極A型的逆戟鯨的分化時間更有33.5萬年���。至於南極A型 , 北大西洋 , 北太平洋居留型及過客型的分類位置還是不明的 , 居留型逆戟鯨及遠洋型逆戟鯨的分化時間則有17萬年之久�����。
在本文為网信彩票論壇精華帖����,由網友兩魏爭雄提供���,僅供大家學習參考���。
編輯����:劉琮瀅
2022-12-03
-

阿根廷發現最大的熊類化石——南美短麵熊
阿根廷大草原早更新世已知最大的熊類——a 南美短麵熊 Arctotherium angustidens���:並討論熊類的體型及飲食趨勢The Largest Known Bear, Arctotherium angustidens, from the Early Pleistocene Pa
阿根廷大草原早更新世已知最大的熊類——a. 南美短麵熊 Arctotherium angustidens��:並討論熊類的體型及飲食趨勢
The Largest Known Bear, Arctotherium angustidens, from the Early Pleistocene Pampean Region of Argentina: With a Discussion of Size and Diet Trends in Bears
Leopoldo H. Soibelzon and Blaine W. Schubert
Abstract
The South American giant short-faced bear (Arctotherium angustidens Gervais and Ameghino, 1880) is one of five described Arctotherium species endemic to South America and it is known for being the earliest, largest, and most carnivorous member of the genus. Here we report an extraordinarily large A. angustidens individual exhumed from Ensenadan sediments (early to middle Pleistocene) at Buenos Aires Province, Argentina. Based on overall size, degree of epiphyseal fusion, and pathologies, this bear was an old-aged male that sustained serious injuries during life. Body mass of the bear is estimated and compared to other ursid species based on a series of allometric equations. To our knowledge, this specimen now represents the largest bear ever recorded. In light of this discovery, we discuss the evolution of body size in Arctotherium (from large-to-small) and compare this to bears that exhibited different evolutionary trajectories. We suggest that the larger size and more carnivorous nature of A. angustidens, compared to later members of the genus, may reflect the relative lack of other large carnivores and abundance of herbivores in South America just after the Great American Biotic Interchange.
翻譯���。���。�����。
南美巨型短麵熊類——a. 南美短麵熊(Arctotherium angustidens����,Gervais and Ameghino, 1880)是南美洲特有的五種南美短麵熊屬(Arctotherium)之一��,它被視為最早�����,已知體型最大而且最食肉性的熊類成員��。
作者描述一個從阿根廷Buenos Aires省Ensenadan沉積物(早~中更新世)掘出的特大a. 南美短麵熊化石���。其化石認為是屬雄性成年體並且生前受過嚴重的傷����。
作者根據異速方程式(allometric equations)估計熊類的體質並比較與其他的物種�����。分析顯示���,這個標本代表有史以來最大的熊屬種��。
作者也討論了南美短麵熊屬體型演變(由大至小)�����,並比較這熊屬的不同進化軌跡及分析其體型和肉食性的衍化 ����。
眼鏡熊亞科(Tremarctinae)化石的地理和時空分布
作者研究的化石標本���。����。����。
左肱骨MLP 35-IX-26-5���,左尺骨MLP 35-IX-26-3和左橈骨MLP 35-IX-26-4
作者描述的最大a. 南美短麵熊化石體型比較����。���。��。
新發現的南美短麵熊(數個標本)化石包括
MLP 35-IX-26-3 左尺骨���, MLP 35-IX-26-4 左橈骨��, MLP 35-IX-26-5 左肱骨���, MLP 35-IX-26-6 右肱骨��, MLP 35-IX-26-7 右橈骨和尺骨
(以上所有化石都是一個個體��,老年雄性����,身前受過傷)�����,
此外還有其幾個其他的大標本個體
MACN 9609���,MACN 5132����,MLP-09-I-5-1���,MLP00-VII-10-1
(這4個用保守估計都至少750kg以上級別��,估計能達到或者接近最大短麵熊的存在F��:AM 25531和UVP015��,如果再算上另外一個論文裏麵的BNMNH 32916和MACN 851的話���,就有7個巨型南美短麵熊標本了)PS����:BNMNH 32916和MACN 851估計也和AM 25531和UVP015差不多大小
和PM 24880(UVP015沒有前肢數據)比較以後可以看出這頭南美巨型短麵熊MLP 35-IX的肢體雖然隻比PM 24880長5—8%����,卻要寬幾乎10—30%����,甚至粗40%的地方也有��,而最大短麵熊UVP015的後肢數據卻大概隻比PM 24880粗10%左右���,長11%����。
UVP015的後肢數據(金閃閃的英雄王在百度發的��,轉一下)
所以可以肯定最大的北美巨型短麵熊UVP015仍然要比著頭新發現南美巨型短麵熊要高一點����,但是粗壯程度則要遜色不少��,體重也要輕於南美巨型短麵眼鏡熊
使用的公式��:
Equations used for body mass prediction. HGDDE = Humerus greatest diameter of distal epiphysis. HMSC = Humerus mid shaft circumference. HGL = Humerus greatest length. RPEGD = Radius proximal epiphysis greatest diameter. ID = equation identification. SEE percent standard error of the estimation. Modified from Soibelzon and Tartarini (2009).
作者認為這種巨型南美短麵熊是肉食性動物(至少偏肉食�����,盡管同屬成員多雜食)����,巨型南美短麵熊的臼齒有磨損和裂痕���,被認識是咀嚼骨頭造成的���,他們有可能和毀滅刃齒虎爭奪獵物��,後來因為南北美交換�����,大型肉食動物數量增加��,競爭加劇���,導致南美短麵熊屬逐漸的變小��。
按公式給最可靠值1600kg確實差不多和中等的白犀牛差不多大了����,但是這個MLP 35-IX-26是老年雄性����,肯定不能拿來代表一般的南美巨型短麵熊的水平�����,其他6個標本基本上都是750-1200kg之間的����。應該也是這個物種的平均大小了��。
脆弱異特龍大的2000-3200kg的還是有不少的����,著頭熊也大概就是中小的成年異特龍水平��。
在本文為网信彩票論壇精華帖���,由網友wingkof提供����,僅供大家學習參考��。
編輯��:劉琮瀅
2022-12-03
-

簡談南北美的史前亞河豚總科屬種
鬱江原白鰭豚(Prolipotes yujiangensis)哈得遜棱河豚(Goniodelphis hudsoni)在淡水豚中白鰭豚科(Lipotidae)和亞河豚總科(Inioidea)關係較密切 , 至於恒河豚總科被認為是喙鯨科(Ziphiidae)及海豚總 科(Delph
鬱江原白鰭豚(Prolipotes yujiangensis)
哈得遜棱河豚(Goniodelphis hudsoni)
在淡水豚中白鰭豚科(Lipotidae)和亞河豚總科(Inioidea)關係較密切 , 至於恒河豚總科被認為是喙鯨科(Ziphiidae)及海豚總 科(Delphinida)的姐妹群����。白鰭豚科(Lipotidae)包括在我國中新世淡水沉積層發現的鬱江原白鰭豚 (Prolipotes yujiangensis) , 晚中新世太平洋東北海岸的海生屬種副拉河豚屬/佩阿波特波鯨屬(Parapontoporia)及剛滅絕的白鰭豚(Lipotes vexillifer)���。鬱江原白鰭豚 (Prolipotes yujiangensis)隻有一小塊吻突殘骸 , 牠被認為和剛滅絕的白鰭豚(Lipotes vexillifer)有密切的關係 , 不過原白鰭豚屬(Prolipotes)位置尚存疑問�����。
亞河豚總科(Inioidea)包括亞河豚科及拉河豚科 , 亞河豚科及拉河豚科耳外骨 , 錘骨 , 前上頜骨及上頜有著共近裔性狀(synapomorphies) , 亞河豚總科(Inioidea)於南美有著不少化石記錄��。
已滅絕的亞河豚科(Iniidae)屬種大多都有極大的體型及較長的吻部 , 化石大多被發現在南美 , 像黃昏亞河豚屬(Hesperoinia) , 蜥河豚屬(Saurocetes)及棱河豚屬(Goniodelphis)化石較殘破 , 牠們過去曾被劃作亞河豚科(Iniidae) , 現被認為可能不屬於亞河豚科(Iniidae)��。
巴塔哥尼亞原亞河豚(Proinia patagonica)被發現在阿根廷早中新世海生組份 , 牠隻有不完整的破碎頭骨及 一塊頸椎化石 , 由於牠的下頜和許多骨骼都嚴重缺失 , 因此有關原亞河豚屬(Proinia)和其它亞河豚亞科屬種的關係存有疑問 , 曾有專家認為巴塔哥尼亞原亞河豚(Proinia patagonica)的骨骼是屬於南鯨屬(Diochotichus)或者和南鯨屬(Diochotichus)有密切關係 , 巴塔哥尼亞原亞河豚(Proinia patagonica) 頭骨和亞河豚屬(Inia)或白鱀豚屬(Lipotes)的有明顯不同特征 , 此外巴塔哥尼亞原亞河豚(Proinia patagonica)有較大的體型���。從化石來看巴塔哥尼亞原亞河豚(Proinia patagonica)和鯊齒鯨屬(Squalodon)較相似且兩者是同時代的 , 如果巴塔哥尼亞原亞河豚(Proinia patagonica)真的是亞河豚亞科成員的話 , 亞河豚類及鯊齒鯨類應該是在早中新世時期已完全分化���。
在加利褔亞San Pablo組發現了近8吋長的下頜聯合 , 下頜聯合有六對齒槽 , 該化石被定種為加利褔亞黃昏鯨(Hesperocetus Californicus) , 根據加利褔亞黃昏鯨(Hesperocetus Californicus)下頜聯合和牙齒形狀而把加利褔亞黃昏鯨(Hesperocetus Californicus)劃歸亞河豚亞科 , 加利褔亞黃昏鯨(Hesperocetus Californicus)牙齒齒冠略呈後彎 , 琺琅質表麵為脊紋所覆蓋�����。
阿根廷蜥海豚(Saurodelphis argentinus)被發現在阿根廷早上新世巴拉那組(Parana formation) , 牠和亞河豚有密切關係 ���。
中新世巴西的本內登等吻河豚(Ischyrorhynchus vanbenedeni)要較現存的亞河豚大出30%且吻部相對的較長 , 本內登等吻河豚(Ischyrorhynchus vanbenedeni)生活在淡水環境����。和其它亞河豚亞科成員的不同 , 等吻河豚屬(Ischyrorhynchus)的顴突置前 , 上頜骨向眶部推延 , 在眼窩上和上顳區旁的上頜骨邊緣向上彎 ,本內登等吻河豚(Ischyrorhynchus vanbenedeni) 一些形態和恒河豚科的相似���。
現存的拉河豚科屬種僅隻有拉河豚(Pontoporia blainvillei)一種 , 拉河豚(Pontoporia blainvillei)的化石被發現在巴西南大河州晚更新世地層���。最古老的拉河豚科化石是秘魯發現的海生屬種短吻海豚屬(Brachydelphis) , 短吻海豚屬(Brachydelphis)生活在中新世中期至晚期 , 該屬近來被認為是所有淡水豚及劍吻海豚科的姐妹群����。
此外在秘魯上新世地層及阿根廷晚中新世地層發現的上新拉河豚屬(Pliopontos)也是海生拉河豚科成員 , 在智利中新世地層也發現了拉河豚科未定種的殘骸 , 在馬裏蘭州晚中新世地層發現了類似上新拉河豚屬(Pliopontos)的部分殘骸 , 此外在佛羅裏達早上新世地層也發現了懷疑是拉河豚科的化石 , 這些化石顯示了拉河豚科的分布已延至北大西洋 , 其中生活在中新世北海一帶 , 拉河豚科的原鼠海豚屬(Protophocaena)有著極廣泛的地理分布��。
阿根廷上新世海洋沉積層發現的直吻擬拉河豚(Pontistes rectifrons)僅隻有顱骨化石發現 , 但是吻端及牙齒缺失 , 直吻擬拉河豚(Pontistes rectifrons)的顱骨和拉河豚屬(Pontoporia)的相似 , 但是直吻擬拉河豚(Pontistes rectifrons)的顱骨要較拉河豚屬(Pontoporia)的大兩倍����。從擬拉河豚屬(Pontistes)的頜骨來看牠的牙齒直徑是拉河豚屬 (Pontoporia)的兩至三倍 , 擬拉河豚屬(Pontistes)牙齒排列並不緊密 , 牠的牙齒數目估計約有53顆至59顆 ���。
至於巴拉那河旁的上新世海生沉積層發現的Pontivaga fischeri僅隻有下頜聯合部分 , 有專家認為Pontivaga fischeri其實是直吻擬拉河豚(Pontistes rectifrons)的同物異名 , 也有專家認為Pontivaga fischeri可能和裂海豚屬(Schizodelphis)存有關連 , Pontivaga fischeri牙齒型態不明���。
近來在加利褔亞聖迭戈更新世地層發現的斯氏尖吻河豚(Stenodelphis sternbergi)隻有一小塊吻突殘骸化石被發現 , 吻突殘骸長約127.5 mm , 該吻部殘骸大約有24顆牙齒����。斯氏尖吻河豚(Stenodelphis sternbergi) 吻突 , 牙齒形狀和拉河豚[Stenodelphis blainvillei \(Pontoporia blainvillei)]的相似 , 斯氏尖吻河豚(Stenodelphis sternbergi)有斧狀的牙根及修長的齒冠 , 牙齒間隔並不緊密 ����。斯氏尖吻河豚(Stenodelphis sternbergi)的牙根和白鱀豚 (Lipotes vexillife)及亞河豚(Inia geoffrensis)的相似 , 但是齒冠琺琅質有所不同���。從斯氏尖吻河豚(Stenodelphis sternbergi)吻突的殘骸來看牠的吻部要較現存的拉河豚[Stenodelphis blainvillei \(Pontoporia blainvillei)]的長����。
北卡羅萊納州早上新世地層發現的貝克奧羅拉鯨(Auroracetus bakerae)化石樣本編號為USNM 534002 , 該頭骨殘骸化石屬幼體 , 保留了完整右鼻骨及鼻骨孔外圍部分 , 上枕骨, 左外枕骨連髁狀突都得以保留 , 可是貝克奧羅拉鯨(Auroracetus bakerae)的吻突及頭蓋骨基部卻沒得以保留���。
古生物學家認為短吻海豚屬(Brachydelphis)屬拉河豚科(Pontoporiidae)最基層的分支 , 其它短吻海豚亞科(Brachydelphinae)成員比如小原鼠海豚(Protophocaena minima)也占據相似的位置��。
拉河豚科(Pontoporiidae)另一支包括擬拉河豚屬(Pontistes) , 上新拉河豚屬(Pliopontos)及拉河豚屬(Pontoporia)則具備兩個特征包括顯現出翼狀骨側葉及具有對稱的顱頂 ���。貝克奧羅拉鯨(Auroracetus bakerae)有對稱的顱頂 , 可是翼狀骨沒被保存下來 , 奧羅拉鯨屬(Auroracetus)看似是屬這個進化支 , 但是上新拉河豚屬(Pliopontos)及拉河豚屬(Pontoporia)前頜骨後邊存有溝漕 , 這是前頜骨的動脈延伸至骨骼後端 , 但在奧羅拉鯨屬(Auroracetus)卻沒發現這特征 , 因此奧羅拉鯨屬(Auroracetus)應該不屬上新拉河豚屬(Pliopontos)及拉河豚屬(Pontoporia)這進化支內 , 但以上隻是屬假設���。如果奧羅拉鯨屬(Auroracetus)和上新拉河豚屬(Pliopontos)及拉河豚屬(Pontoporia)關係密切的話 , 那麽北美東岸的化石發現變得相當平常 , 現存的拉河豚屬(Pontoporia)在南美東岸出沒 , 牠們的共同先祖沿著淺海岸向北擴散進入北美這理論就變得相當合理了���。假設奧羅拉鯨屬(Auroracetus)和早上新世秘魯的上新拉河豚屬 (Pliopontos)關係較接近的話 , 牠們的共同先祖一定要橫過中美最後到達巴拿馬地峽 ���。另一點值得注意的是 , 在北美Lee Creek礦山和秘魯Sud-Sacaco都同樣發現了喙鯨科的利隆喙鯨屬(Ninoziphius)化石 , 這是一個早上新世時期鯨類於兩地擴散的成功例子 , 不過還需要更多拉河豚科的化石來證實拉河豚科屬種之間的關係���。
在本文為网信彩票論壇精華帖����,由網友耽羅山水提供���,僅供大家學習參考���。
編輯����:劉琮瀅
2022-12-03
網信彩票
